


1Singapore Immunology Network (SIgN), A*STAR (Agency for Science, Technology and Research), Biopolis, 138648 Singapore., 2National Skin Centre, 3School of Biological Sciences, 4Department of Microbiology and Immunology, Yong Loo Lin School of Medicine, and 5Lee Kong Chian School of Medicine, Nanyang Technological University, Singapore, Singapore. E-mail: Ng_Lai_Guan@immunol.a-star.edu.sg
#Both authors contributed equally to this work.
Accepted Feb 20, 2018; Epub ahead of print Feb 28, 2018
Vasculitis is a state of inflammation of the blood vessel walls. Different types of vasculitides are categorized based on the size of vessels involved in inflammation, as well as their aetiology, pathogenesis and type of inflammation (1). The hallmark symptoms of Henoch-Schönlein purpura (HSP) are purpura, arthralgia and gastrointestinal involvement, due to systemic inflammation. Renal involvement is present in 45–85% of patients with HSP, manifesting with haematuria or proteinuria (2), and is probably due to the deposition of immune complexes (ICs) in the mesangium of the kidney. In some cases, renal involvement progresses to end-stage renal failure. We demonstrate here that it is possible to interrogate skin biopsies to screen for potential candidate genes that may be involved in the pathogenesis of the disease.
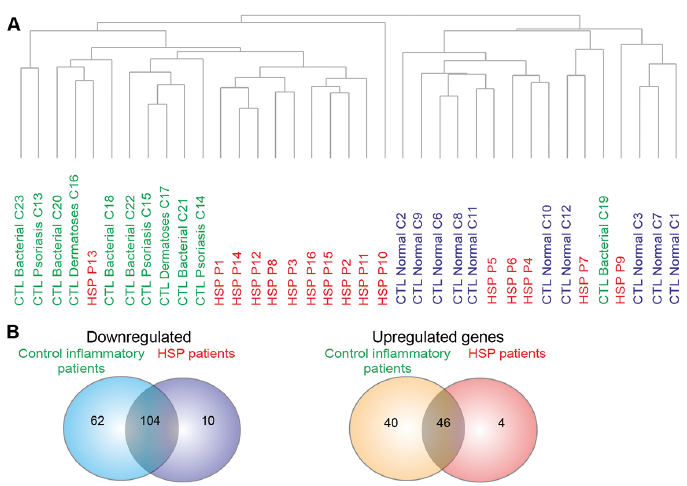
Nanostring analysis was run on RNA extracted from skin biopsies from 3 groups of samples: normal skin (n = 10), control patients with different inflammatory skin diseases with known neutrophil infiltration (n = 11), and patients with HSP (n = 16) with similar sex ratio and age ranges. The resultant data was normalized using housekeeping genes as a reference (Fig. S1). Unsupervised clustering was then carried out using genes with expression levels above background to determine whether samples from different patient categories can be distinguished from each other in an objective manner. The dendrogram reveals the heterogeneity of patients with HSP, as 62.5% of the patients with HSP are distinct from individuals with normal skin and control inflammatory patients (HSP-inflamed), while 31.25% of patients with HSP cluster together with individuals with normal skin (HSP-normal) (Fig. 1A).
Analysis of variance (ANOVA) analysis was first used to identify differentially expressed genes (DEGs) amongst individuals with normal skin, inflammatory control patients and all patients with HSP, followed by post-hoc t-tests. All genes were expressed as fold change from normal skin. Of the 575 genes with expression levels above the background in patients with HSP, 114 genes were downregulated, and 50 were upregulated. However, after eliminating genes with similar changes in control inflammatory patients, only 10 of these genes were downregulated, whereas 4 were upregulated (Fig. 1B). These 14 genes thus probably represent those linked with the pathology of HSP.
Fig. 1. Nanostring analysis reveals differentially expressed genes and heterogeneity in patients with Henoch–Schönlein purpura (HSP). (A) Dendrogram of samples based on the gene expression levels (log10 normalized counts) above background. Samples of individuals with normal skin (blue); control inflammatory patients (green) and patients with HSP (red). Dendrogram was computed using hierarchical clustering of the log10 normalized counts using Euclidean distance and average linkage. (B) Venn diagram illustrating the number of upregulated and downregulated genes for both the control inflammatory patients and patients with HSP.
Amongst the DEGs, 3 out of 4 upregulated genes are known to be associated with other forms of small vessel vasculitides (Table SI). For instance, CCL26 is known to be upregulated in patients with eosinophilic granulomatosis with polyangiitis (EGPA) (3). Six out of 10 of the downregulated genes are known to be linked with other forms of vasculitides. Cytotoxic T-lymphocyte associated protein-4 (CTLA-4) is a T-cell inhibitory molecule expressed on activated T cells, and polymorphisms in the CTLA-4 gene are associated with ANCA-associated vasculitis patients (4). In our screen, 3 genes: STAT-6, arginase-2 and MALT-1 are not previously known to be involved in vasculitis or HSP. Ingenuity pathway analysis of the entire HSP cohort was used to provide a biological context to interpret the Nanostring data. This revealed that the canonical pathways IL-6, hepatocyte growth factor (HGF) and IL-10 are altered in patients with HSP (Complete data can be obtained from the corresponding author).
As there was demonstrable heterogeneity amongst patients with HSP, post-hoc t-tests were carried out to detect DEGs between patients with HSP clustering with individuals with normal skin (HSP-normal) and patients with HSP in a separate cluster (HSP-inflamed). DEGs between the 2 clusters made up approximately 30% of the gene panel. Notably, IPA of the 2 HSP clusters revealed that the top 10 functional pathways affected were related to the expansion of different T-cell subsets and development of dendritic cells (2). Interestingly, these genes were comparatively downregulated in the HSP inflamed cluster compared with the HSP normal cluster (Fig. S2). This suggested that there could be downregulation of genes involved in inflammation.
Notably, classical pro-inflammatory cytokines and genes linked with neutrophil recruitment were significantly lower in the HSP-inflamed cluster compared with normal skin (Fig. S3). A closer look at the downregulated genes in HSP-inflamed cluster reveals that approximately 40% (51 out of 130) genes in our panel known to be involved in the gene ontology (GO) classification of inflammatory response (GO: 0006954) are downregulated compared with approximately 10% (14 out of 130) that was upregulated. This trend is similar for genes positively regulating cytokine production (GO: 0001819) (Fig. S4), and control inflammatory patients demonstrate a similar trend.
This study demonstrated the viability of using formalin-fixed paraffin-embedded (FFPE) archival skin tissues to perform mRNA analysis. Conventionally, archival histological specimens cannot be used for mRNA studies, as degradation of RNA precludes RNA sequencing. NanoString eliminates the need for RNA amplification, as it utilizes tagged capture probes that hybridize to target molecules, which are then counted. This is one of the first studies demonstrating mRNA analysis in archival skin specimens through the use of NanoString (5, 6).
We chose patients with inflammatory conditions where there is known neutrophil infiltration (Appendix S1) as the control inflammatory group, to aid in distinguishing DEGs associated with HSP pathology from genes linked with general neutrophil infiltration. However, it should be noted that the heterogeneity in the control inflammatory patients is also a limitation of our study, since we may miss the discovery of other less strongly associated DEGs due to the increased variability.
DEGs identified in patients with HSP, but not in the control inflammatory group, are linked with regulation of T-cell activation, such as CTLA-4, CXCR-6, DPP-4 and IL-21R, which is expected, since T-cell help is necessary for B-cell production of antibodies. Genes not known to be associated with HSP or vasculitis are involved in immune activation in different ways. STAT-6 is a transcription factor for IL-3 and IL-4 production; arginase-2 is thought to play an anti-inflammatory role in macrophages and MALT-1 is involved in NF-κB activation. The full list of genes that are significantly changed in our study can be requested from the authors. Notably, the IL-6 and IL-10 pathways are involved as they are important for B-cell activation and maturation.
Interestingly, initial clustering analysis revealed heterogeneity in the HSP patient cohort. The clustering result is independent of confounding factors, such as RNA amount, RNA integrity number (RIN) and sample age. Ingenuity Pathway Analysis (IPA) of the clusters (complete data can be obtained from the corresponding author) showed differences in many T-cell-related pathways, such as expansion of Th17 cells and T lymphocytes, and inhibition of effector T lymphocytes. This could be due to differential levels of T cell activation, as majority of these genes for the HSP-inflamed group was downregulated. The list of DEGs between HSP-inflamed and HSP-normal clusters can be requested from the authors.
It is likely that the downregulation of genes linked with inflammatory responses is due to suppression of these genes, and the results of the current study indicate that care should be taken when interpreting the downregulation of genes in screens such as NanoString analysis.
In summary, our data correlates with existing data on vasculitis, and pharmacological targeting of the identified pathways could be useful in HSP treatment. On the other hand, some of the genes identified in this study such as STAT-6, arginase-2 and MALT-1 have not been described in relation to vasculitis, and further investigations can be carried out for these genes to determine if they are potential biomarkers for HSP. Our results also reveal heterogeneity within the patients with HSP in their responses, and it would be interesting to further investigate the causes of the different responses in these 2 groups of patients.
Funding for this study was provided with a grant from the National Healthcare Group.
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize