


Ingrid Asp Psoriasis Research Center, Department of Clinical and Experimental Medicine, Linköping University, Linköping, Sweden
Psoriasin, which is highly expressed in psoriasis, is encoded by a gene located within the epidermal differentiation complex. The aim of this study was to investigate the effect of endogenous psoriasin on disturbed keratinocyte differentiation in psoriasis. Immunohistochemical staining revealed a gradient of psoriasin expression in the psoriatic epidermis with highest expression in the suprabasal, differentiated layers. Induction of keratinocyte differentiation caused concurrent expression of psoriasin and the differentiation marker involucrin. The differentiation-induced psoriasin expression was found to be mediated by the protein kinase C pathway. The downregulation of psoriasin expression by small interfering RNA revealed that psoriasin mediates the expression of involucrin, desmoglein 1, transglutaminase 1 and CD24 in normal differentiation. The lentivirus-mediated overexpression of psoriasin, mimicking the psoriatic milieu, gave rise to an altered regulation of differentiation genes and an expression pattern reminiscent of that in psoriatic epidermis. These findings suggest that psoriasin contributes to the dysregulated differentiation process in the psoriasis epidermis.
Key words: psoriasin; S100A7; differentiation; keratinocyte; psoriasis.
Accepted Dec 12, 2016; Epub ahead of print Dec 13, 2016
Acta Derm Venereol 2017; 97: xx–xx.
Corr: Charlotta Enerbäck, Ingrid Asp Psoriasis Research Center, Department of Clinical and Experimental Medicine, Linköping University, SE-581 85 Linköping, Sweden. E-mail: charlotta.enerback@liu.se
To establish the skin barrier and the epidermal structure, keratinocytes undergo a tightly regulated process of differentiation. In this process, the keratinocytes progress from the basal layer of the epidermis through distinct stages to form the spinous, granular and cornified layers. Each of these layers is characterized by specific features in terms of gene expression, function and cellular mor-phology (1).
In psoriasis, the differentiation process of the epidermal cells is disturbed. The keratinocytes in active psoriatic lesions display an altered sequence and altered expression levels of the differentiation markers (2). However, the mediators that are responsible for the dysregulated differentiation of psoriatic keratinocytes have not been identified.
Psoriasin (S100A7) was first identified for its overexpression in psoriatic keratinocytes (3), where it was found to be located to the nucleus and the cytoplasm (4). Later studies have demonstrated secretion of psoriasin from epithelial cells (5). Intracellular psoriasin has been found to interact with various signalling proteins, including the nuclear factor (NF)-kB regulator Jab1, while secreted psoriasin functions as an antimicrobial peptide and has the ability to induce chemotaxis of neutrophils and T cells (6). Psoriasin is strongly induced by interleukin (IL)-22 and by the combination of IL-17 and tumour necrosis factor (TNF)-α, which emphasizes the role of psoriasin in the pathogenesis of psoriasis (7). We have shown previously that extracellular psoriasin binds to the receptor for advanced glycation end products (RAGE), promoting endothelial cell proliferation (8). In addition to the low expression in normal skin and the high expression in the psoriatic epidermis, psoriasin is also expressed in conditions displaying abnormal differentiation. In a spectrum of conditions of dysregulated keratinocyte differentiation, we have previously demonstrated strong psoriasin expression in differentiated squamous cell carcinoma and a lack of expression in undifferentiated basalioma (9). This pattern of expression suggests an involvement in epithelial differentiation. In addition, the gene that encodes psoriasin is positioned within the epidermal differentiation complex (EDC) (10–12), a gene cluster that contains several genes of importance in the terminal differentiation of the human epidermis (10, 11).
The high expression level in psoriatic epidermis in conjunction with the disturbed differentiation process suggests that psoriasin is involved in keratinocyte differentiation. Previous studies of the role of psoriasin in keratinocyte differentiation have focused on the effects of extracellular psoriasin, but as the psoriasin in psoriasis is keratinocyte-derived, it is imperative to consider the effects of psoriasin as an endogenously produced intracellular mediator. The aim of this study was therefore to investigate the effect of keratinocyte-derived, intracellular psoriasin on keratinocyte differentiation.
Neonatal human epidermal keratinocytes (HEKn; Life Technologies, Carlsbad, CA, USA), were cultured in EpiLife medium supplemented with 1% EpiLife defined growth supplement (EDGS), 0.06 μM CaCl2 (Gibco, Paisley, UK) and 1% penicillin/streptomycin (PAA Laboratories, Pasching, Austria). The keratinocyte cell line NTERT-2G was grown in keratinocyte serum-free medium (KFSM, Gibco) as described previously (13). Extracellular treat-ment with psoriasin (Abnova, Taipei, Taiwan) was conducted with 0.15 µg/ml or 1 µg/ml for 72 h.
Skin punch biopsies were obtained from healthy controls and lesional psoriasis skin and were fixed and paraffin-embedded. The sections were deparaffinized in Histolab-clear (Histolab Products, Gothenburg, Sweden) and rehydrated in ethanol. Heat-induced antigen retrieval was performed in citrate (pH = 6) antigen retrieval buffer (DAKO, Glostrup, Denmark). Endogenous peroxidases were blocked with 3% hydrogen peroxide, and unspecific protein binding was inhibited with 5% bovine serum albumin. Antibody binding with primary mouse anti-human psoriasin antibody (1:400; Imgenex, San Diego, CA, USA) was carried out overnight at 4°C. Following incubation, DAKO EnVision anti-mouse HRP-conjugated polymer was applied, and antigen-antibody complexes were visualized by DAB-staining (PolySciences, Eppelheim, Germany). Cell nuclei were counterstained with haematoxylin (Invitrogen, Paisley, UK).
Keratinocyte differentiation was induced by treating HEKn with CaCl2 or 12-O-tetradecanoylphorbol-13-acetate (TPA) or by culturing HEKn in suspension or confluence. Differentiation was confirmed by measuring involucrin. For differentiation with CaCl2 or TPA, HEKn were treated with 2 mM CaCl2 (Sigma-Aldrich, St Louis, MO, USA) or 1 μM TPA (Acros Organics, Geel, Belgium) for 48 h. For suspension culture, 24-well plates were coated with poly-HEMA. The cells were seeded in complete EpiLife and harvested after 24 or 48 h. For differentiation in confluence, cells were seeded at near-confluence in complete EpiLife medium. Upon cell confluence, the culture medium was substituted for EDGS-free EpiLife medium and cells were grown without any additional change of medium for 5 or 7 days.
To investigate the participation of the protein kinase C (PKC) pathway, HEKn were pre-treated with the PKC-inhibitor bisindolylmaleimide I (4 μM, Cell Signaling, Danvers, MA, USA), or the PKC-δ inhibitor rottlerin (4 μM, Cell Signaling) for 30 min followed by treatment with CaCl2. An equal volume of dimethyl sulfoxide (DMSO) was included in non-inhibitor wells as vehicle control.
To down-regulate psoriasin expression, HEKn were transfected with psoriasin-siRNA (Pso-siRNA) or a validated non-targeting siRNA (C-siRNA) (Santa Cruz Biotechnology, Santa Cruz, CA, USA). Following transfection, the cells were treated with 2 mM CaCl2 for 48 h.
Apoptosis was analysed by the FITC Annexin V Apoptosis Detection Kit I (BD Biosciences, San Jose, CA, USA), staining cells with Annexin V-FITC and propidium iodide (PI). Cell staining was analysed using a Gallios flow cytometer and Kaluza analysis software (Beckman Coulter, Brea, CA, USA). Annexin V–PI– cells were considered to be viable, Annexin V+PI– cells apoptotic and Annexin V+PI+ cells post-apoptotic necrotic.
Human NTERT keratinocytes with a sustained overexpression of psoriasin were generated as described previously (14). Briefly, total RNA was extracted from psoriatic skin biopsies and cDNA was synthesized. Psoriasin (NM_002963) was amplified by PCR and subsequently cloned into the lentiviral expression vectors pLenti6.3/TO/V5-DEST and used to produce infectious lentivirus particles.
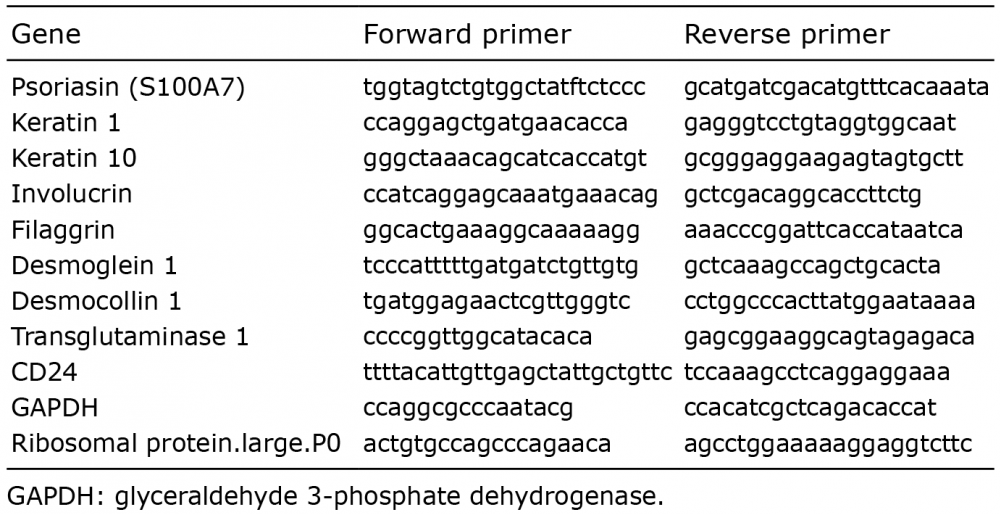
The RNeasy mini kit (Qiagen, Hilden, Germany) was used to extract RNA. cDNA was synthesized using the Maxima First Strand cDNA Synthesis Kit (Fermentas, Vilnius, Lithuania). qPCR was performed with the SYBR green (Applied Biosystems, Foster city, CA, USA) on a real-time 7500HT or 7900HT PCR (Applied Biosystems). Primers for psoriasin and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) were designed as described (15–18). Primers for ribosomal protein large p0 (RPLP0), cytokeratin (K)1, K10, involucrin, filaggrin, desmoglein (DSG)-1, desmocollin (DSC)-1, transglutaminase (TGM)-1 and CD24 were designed using the Primer Express Software version 3.0 (Applied Biosystems) (Table I). Expression data were normalized to RPLP0 or GAPDH as indicated, using the comparative Ct (2–ΔΔCt) method.
Table I. Primer sequences used for quantitative real-time PCR
Keratinocytes were fixed in 4% paraformaldehyde and permeabilized with 0.1% saponin. The cells were incubated overnight at 4°C with monoclonal mouse anti-human anti-psoriasin (Abcam, Cambridge, UK), followed by incubation with secondary goat anti-mouse AlexaFluor555 (Molecular Probes, Eugene, OR, USA). Nuclei were counterstained with DAPI (4’,6-diamidino-2-phenylindole). Staining was evaluated in an AxioVert.A1 Microscope (Zeiss, Oberkochen, Germany). Negative controls were obtained by omitting the primary antibody and only incubate the sections with secondary antibody. These controls displayed no staining.
To determine the co-occurrence of psoriasin, S100A8, S100A9 and S100A12 in psoriasis, skin punch biopsies, 4-mm in diameter, were obtained from age- and sex-matched patients with psoriasis and from healthy controls. A total of 9 biopsies, consisting of 3 involved (PP), 3 uninvolved (PN) and 3 normal (NN) skin biopsies, were obtained. The epidermis was detached by soaking the biopsy in 3.8% sterile ammonium thiocyanate in phosphate-buffered saline (PBS) for 30 min, as previously described, a method that does not induce any changes in gene expression (18). Following incubation, the epidermis was removed and RNA was isolated immediately.
RNA extracted from 3 psoriasin-overexpressing or control N-TERTs or from PP, PN and NN epidermis was used to generate hybridization probes using the GeneChip® WT PLUS Reagent kit according to the manufacturer’s instructions. Hybridization to GeneChip® human transcriptome arrays 2.0 was performed in a GeneChip® hybridization oven, followed by washing and staining in a GeneChip® Fluidics Station 450 and scanning in a GeneChip® Scanner 3000 7G system. Expression Console software (v1.2.1) was used to verify the quality controls. All GeneChip® products were from Affymetrix (Santa Clara, CA, USA). CEL-files generated by the Affymetrix AGCC program were analysed using Genespring GX software (v13.0; Agilent Technologies, Santa Clara, CA, USA). A Standard Robust Multi-Array Average (RMA) normalization and background correction was performed. The probe sets were filtered on expression with a 20th percentile cut-off. The fold change was computed by the GeneSpring software. On the probe sets that remained after the filtering, a 1.5-fold change was applied as a cut-off for up- and down-regulated genes. To determine the co-expression of S100 proteins, a heatmap was constructed in R (version 3.2.4) and clustering was performed based on normalized data.
The data are presented as the mean ± standard error of mean (SEM), and were compared using paired Student’s t-test or Wilcoxon signed-rank test. Analyses were performed using GraphPad Prism 6 software (GraphPad software, La Jolla, CA, USA). A value of p < 0.05 was considered statistically significant.
Psoriasin was originally identified as a protein highly expressed in lesional psoriatic skin (3). To describe the differential expression pattern between patients with psoriasis and healthy controls, we performed immunohisto-chemistry and obtained a sparse psoriasin expression in normal skin (Fig. 1a) that was not noticeably different from the negative control (Fig. 1c). In the lesional psoriatic skin, we observed a pronounced staining (Fig. 1b) with a tendency for a psoriasin gradient, ranging from weak expression in the basal cell layer to intense expression in the more differentiated layers of the epidermis. This pattern of psoriasin expression suggests the involvement of psoriasin in later stages of keratinocyte differentiation.
Fig. 1. Psoriasin is expressed as a gradient in the psoriatic skin. Immunohistochemical staining of (a) normal and (b) psoriatic skin demonstrates the epidermal expression of psoriasin in psoriatic skin, but not in normal, healthy skin. Psoriasin displays a substantially higher staining intensity in the upper, more differentiated layers of the psoriatic skin. The weak expression in the basal cells and the intense expression in cells of the suprabasal layers present a gradient of psoriasin expression. Primary antibody was applied at a dilution of 1:400, and (c) panel displays the negative control, which was obtained by omitting the primary antibody. The figures are representative of 5 experiments. × 20 magnification was used.
An effect of extracellular, exogenous psoriasin on keratinocyte differentiation has been previously suggested by Hattori et al. (19) and by Son et al. (20). To examine the effects in our culture system, HEKn were treated with psoriasin and the expression of involucrin and psoriasin were investigated. It was found that exogenous psoriasin increased involucrin, but also that it caused up-regulated psoriasin expression, suggesting that extracellular psoriasin may induce intracellular production in an autocrine fashion (Fig. 2a). We did not observe any change in cell morphology upon psoriasin treatment, suggesting that extracellular psoriasin may be unable in itself to induce differentiation.
Fig. 2. The endogenous psoriasin expression upon differentiation is regulated by the protein kinase C (PKC) pathway. (a) To determine the effects of exogenous psoriasin, neonatal human epidermal keratinocytes (HEKn) were treated with psoriasin at concentrations of 0.15 µg/ml and 1 µg/ml for 72 h. (b) To examine the effect of endogenous psoriasin expression for differentiation, HEKn were treated with 2 mM CaCl2 for 48 h or 1 μM tetradecanoylphorbol-13-acetate (TPA) for 48 h or cultured in suspension or in confluence. Calcium and TPA treatment both induced the expression of psoriasin and involucrin. Psoriasin and involucrin expression was also induced in cells cultured in suspension or in confluence. Expression levels are shown as relative fold induction compared with untreated control, using RPLP0 as the internal control. To further study the role of PKC, HEKn were pre-treated with 4 μM of (c) the general PKC-inhibitor bisindolylmaleimide I, (d) the PKC-δ-specific inhibitor rottlerin, or vehicle DMSO for 30 min, followed by treatment with 2 mM CaCl2 for 48 h. CaCl2 induced mRNA expression of psoriasin and involucrin, was reduced by the inhibition of PKC. Expression levels are shown as relative fold induction compared with untreated control using RPLP0 as the internal control. Data are displayed as mean ± SEM, n = 6; *p < 0.05, **p < 0.01.
To evaluate whether a relationship between psoriasin and differentiation rather than merely a co-occurrence of gradients exists, psoriasin expression was evaluated in cells exposed to differentiation-inducing conditions. Keratinocytes were treated with calcium, TPA or were cultured in suspension or confluence, all of which are established differentiation methods (21–24). All these conditions induced the expression of both psoriasin and involucrin (Fig. 2b). In conclusion, we found that psoriasin is endogenously produced in keratinocytes in response to differentiation.
PKC has been implicated in the regulation of keratinocyte differentiation (25) and inhibitors of the PKC-δ pathway block the calcium-induced expression of involucrin (26). To further evaluate the role of endogenous psoriasin in keratinocyte differentiation we investigated the effect of PKC inhibitors in differentiated keratinocytes. Keratinocytes with intact PKC signalling displayed induced expression of psoriasin and involucrin on calcium treatment. Treatment with the general PKC inhibitor bisindolylmaleimide (Fig. 2c) or with rottlerin (Fig. 2d), a specific PKC-δ inhibitor, markedly reduced the expression both of psoriasin and involucrin. These results demonstrate that PKC-δ is involved in the enhanced psoriasin expression in keratinocyte differentiation.
To determine whether psoriasin is part of the keratinocyte differentiation pathway, or merely simultaneously induced, cells were transfected with siRNA. The successful down-regulation of psoriasin expression using siRNA was confirmed by qPCR (Fig. 3a). The differentiation process was induced using calcium, which triggered the expression of the differentiation markers filaggrin, DSG-1, DSC-1, TGM-1 and CD24 (Fig. 3b–g). Furthermore, calcium treatment induced the expression of K1 and K10 (data not shown). The down-regulation of psoriasin resulted in a significantly diminished expression of involucrin (Fig. 3b), DSG-1 (Fig 3d), TGM-1 (Fig. 3f) and CD24 (Fig. 3g). The expression of filaggrin was decreased, but did not reach statistical significance (Fig. 3c). No effect of the down-regulation was seen on the differentiation markers DSC-1 (Fig. 3e), K1 or K10 (data not shown), suggesting that the importance of psoriasin on the differentiation process may vary in different stages of differentiation. This suggests that endogenous psoriasin is an important mediator during the later stages of keratinocyte differentiation.
Fig. 3. The down-regulation of psoriasin expression by siRNA suppresses normal keratinocyte differentiation. Neonatal human epidermal keratinocytes (HEKn) were transfected with psoriasin-siRNA (Pso-siRNA) or controlsiRNA (C-siRNA) followed by treatment with 2 mM CaCl2 for 48 h. The mRNA expression of psoriasin, involucrin, filaggrin, desmoglein (DSG)-1, desmocollin (DSC)-1, transglutaminase (TGM)-1 and CD24, analysed with quantitative real-time PCR (qPCR), was induced by calcium treatment (a-g). (b) Involucrin, (d) DSG-1, (f) TGM-1 and (g) CD24 mRNA expression was decreased by down-regulation of psoriasin. (c) The reduced expression of filaggrin did not reach statistical significance. (e) No reduction of DSC-1 mRNA expression was demonstrated by down-regulation of psoriasin. Expression levels are shown as relative fold induction compared with untreated control using glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as the internal control. Data are displayed as mean ± standard error of the mean (SEM), n = 6; *p < 0.05.
The down-regulation of psoriasin expression by siRNA did not alter cellular morphology and did not affect cell proliferation compared with control cells (data not shown). Likewise, we did not see any difference in either apoptosis or necrosis in siRNA-treated keratinocytes compared with control cells (data not shown). This supports the results we observed with the extracellular psoriasin, again indicating that psoriasin does not, on its own, induce differentiation, nor does it affect apoptosis. It also confirms that the effects we see are not due to increased cell toxicity.
To evaluate the effect of a high expression of psoriasin, as seen in psoriasis, keratinocytes with a sustained overexpression were generated. Immunofluorescence demonstrated a strong staining in the vector-transfected keratinocytes, but no staining in non-transfected cells (Fig. 4a). The transfection did not affect the cell morpho-logy (Fig. 4b). Using microarray, we found a profound effect of psoriasin overexpression on mRNA transcripts related to differentiation. Interestingly, among the top 5 most down-regulated genes in the microarray of psoriasin-overexpressing keratinocytes compared with control cells, 4 are associated with differentiation, namely DSG-1, CASP14, K10 and DSC-1 (Table II). Among the top 15 most down-regulated genes, we observed the differentiation-regulating genes TGM-1, K1, FOXN1 and the SPRR1A gene. Among the up-regulated genes in the microarray in the psoriasin-overexpressing cells, the gene coding for involucrin was found to be the 11th most upregulated gene. Furthermore, the overexpression of psoriasin was also found to cause an up-regulation of IL1B and K16 and a down-regulation of CXCL14.
Fig. 4. Keratinocytes overexpressing psoriasin display signs of dysregulated differentiation, and psoriasin (S100A7), S100A8, S100A9, S100A12 and S100A7A (S100A15) are co-expressed. NTERT keratinocytes were vector-transfected to generate a sustained overexpression of psoriasin (psoriasin-NTERTs) and were compared with control NTERT keratinocytes lacking the overexpression (NT1). Psoriasin protein expression was verified using: (a) immunofluorescence compared with control keratinocytes. Alexa555-conjugated secondary antibody (red) was used to visualize psoriasin expression and DAPI (4’,6-diamidino-2-phenylindole) (blue) was used for nuclear staining. The images are representative of 4 experiments. Panel (b) displays the cell morphology of the psoriasin-overexpressing NTERT keratinocytes and the NTERT control keratinocytes. The images are representative of 4 experiments. Panel (c) demonstrates the proliferation of the psoriasin-overexpressing NTERTs and the control cells. Data are displayed as mean±standard error of the mean (SEM), n = 4; ***p ≤ 0.001. (d) Heatmaps illustrating the clustering of S100-proteins in lesional psoriatic (PP), non-lesional psoriatic (PN) and healthy keratinocytes (NN), n = 3.
Table II. The top 15 most up-regulated and down-regulated differentially expressed genes in psoriasin-overexpressing
NTERTs compared with control NTERTs
These results support that psoriasin has a dysregulating function on keratinocyte differentiation. Keratinocytes that overexpressed psoriasin also exhibited a significantly lower proliferation than control keratinocytes (Fig. 4c), strengthening that these cells undergo differentiation with the accompanying decrease in proliferation.
Psoriasin is part of a family of S100-proteins with high sequence homology. To determine whether other proteins in this family share the expression pattern of psoriasin in psoriasis, we constructed a heatmap, comparing the expression of S100-proteins between lesional psoriasis, non-lesional psoriasis and healthy epidermis. We found a strong co-clustering of psoriasin and S100A8, S100A9 and S100A12, suggesting a co-regulation of these proteins (Fig. 4d).
Keratinocyte differentiation is coordinated by an intricate sequence of gene activation (27). Several markers of differentiation have been identified in epidermal keratinocytes, with involucrin being the most established. The expression of involucrin commences in the upper spinous layer and continues through the granular layer (28) in a similar localization pattern as TGM-1 (29). In contrast, K1 and K10 are considered markers of early differentiation (30). The stratum corneum is marked by the expression of filaggrin (31). CD24 is most prominent in differentiated mammary epithelium (32, 33), but has also been detected in the upper, more differentiated layers of human epidermis (34).
Psoriasis is characterized by an epidermal thickening with an absence of the granular layer and a thickened and irregular cornified layer. Some differentiation markers, including involucrin and TGM, appear prematurely in the lower layers of the psoriatic epidermis, while other markers, such as filaggrin, are either absent or found in the cornified layer. The expression levels of several markers are abnormal, with some markers, including involucrin, demonstrating an overexpression (2, 35), and others, such as K1 and K10, being down-regulated (36).
Previous studies that have attempted to elucidate the role of psoriasin in keratinocyte differentiation have focused on the effects of extracellular, exogenous psoriasin, with conflicting results. While Hattori et al. (19) demonstrated up-regulation of differentiation markers after psoriasin treatment, Son et al. (20) suggested that exogenous psoriasin may negatively regulate differentiation. The findings by Hattori et al. are in line with the increase in involucrin that we observe upon psoriasin treatment. Applying several methods of induction of differentiation, we found that all these methods induced the expression of involucrin and psoriasin, supporting that psoriasin is involved in the differentiation process. PKC is considered a key pathway of epidermal differentiation (26, 37). PKC activation leads to an up-regulation of S100A8 and S100A9 (38), while inhibition of PKC blocks calcium-dependent involucrin expression (39). We reveal an association between psoriasin and involucrin by the shared PKC regulation.
To investigate the role of intracellular psoriasin in normal keratinocyte differentiation, we performed a knock-down of psoriasin expression. This knock-down revealed that in the normal differentiation process, psoriasin acts as a mediator for involucrin, DSG-1, TGM-1 and CD24 expression. This is the first time psoriasin is found to regulate these markers. The induction of involucrin upon differentiation has been described previously, but the mediating role of psoriasin in its expression is a novel finding. Similarly, psoriasin has been suggested to be a substrate for transglutaminases (40), but is here found to also regulate them. We have previously reported that CD24 expression is regulated by psoriasin in mammary epithelial cells (33), and now demonstrate a similar mechanism in keratinocytes. The induction of tight cell-cell interaction by high calcium levels through the formation of desmosomes has been described previously (41, 42). Consistent with this, we found a calcium-induced increase in DSG-1 expression that was abolished by the down-regulation of psoriasin. These results suggest that the expression of psoriasin is correlated with markers of differentiation in normal skin, although our findings do not necessarily indicate the involvement specifically in late differentiation.
While psoriasin is only weakly present in normal skin, it is markedly up-regulated in psoriasis. The effects of psoriasin on differentiation are therefore likely to differ from those in normal skin. To mimic this up-regulation, we generated keratinocytes with a sustained overexpression of psoriasin. We found that the sustained intracellular overexpression of psoriasin regulates several markers of differentiation, manifesting in up-regulation of involucrin, and down-regulation of DSG-1, caspase-14, K10, DSC-1, FOXN1, TGM-1 and K1. These changes in gene expression reflect the pattern seen in psoriasis epidermis, where involucrin is up-regulated and caspase-14, K10 and K1 down-regulated (43, 44). Furthermore, the overexpression of psoriasin was also found to cause up-regulation of the cytokine IL1B, which is locally increased in psoriasis (45) and down-regulation of the chemokine ligand CXCL14, which is decreased in psoriasis (46). We also identified overexpression of K16, in the psoriasin overexpressing cells, consistent with what has been described in psoriasis (44).
Our heatmap, based on the transcriptome of psoriatic epidermis and control skin, reveals a prominent co-expression of the homologue S100 proteins psoriasin, S100A8, S100A9 and S100A12, suggesting a co-regulation and a putative redundancy. They may also synergize in their effect to promote differentiation. Interestingly, S100A8 and S100A9 have been implicated in keratinocyte differentiation (47). The MAPK signalling pathway has been implicated in the regulation of S100A8/9 (48). Sphingosine kinases regulate NOX2 activity via p38 MAPK-dependent translocation of S100A8/A9 (48). A recent publication showed that up-regulation of psoriasin on TLR3 activation is mediated by p38 (49). This suggests an interesting overlap of pathways between these S100 proteins.
In conclusion, this study demonstrates that psoriasin affects keratinocyte differentiation in a concentration- dependent manner. In the normal differentiation process, psoriasin regulates the expression of several intermediate and late differentiation markers. However, when overexpressed at pathologically relevant levels, the effects are different than those seen upon induction in normal skin, yielding an expression pattern of differentiation markers that resembles that seen in lesional psoriatic skin. While psoriasin does not on its own induce differentiation, it is clear that it acts as a modulator. Instead of merely acting as a positive or negative regulator of differentiation, psoriasin appears to skew the process. These findings suggest that psoriasin is a prominent contributor to the dysregulated differentiation process that has been established in the psoriatic epidermis.
The authors would like to thank Stefan Stoll for help with generating the psoriasin-overexpressing NTERTs.
This research was funded by the Ingrid Asp Foundation, the Welander Foundation, the Swedish psoriasis association and the Medical Research Council.
The authors declare no conflicts of interest.


 Click to show fullsize
Click to show fullsize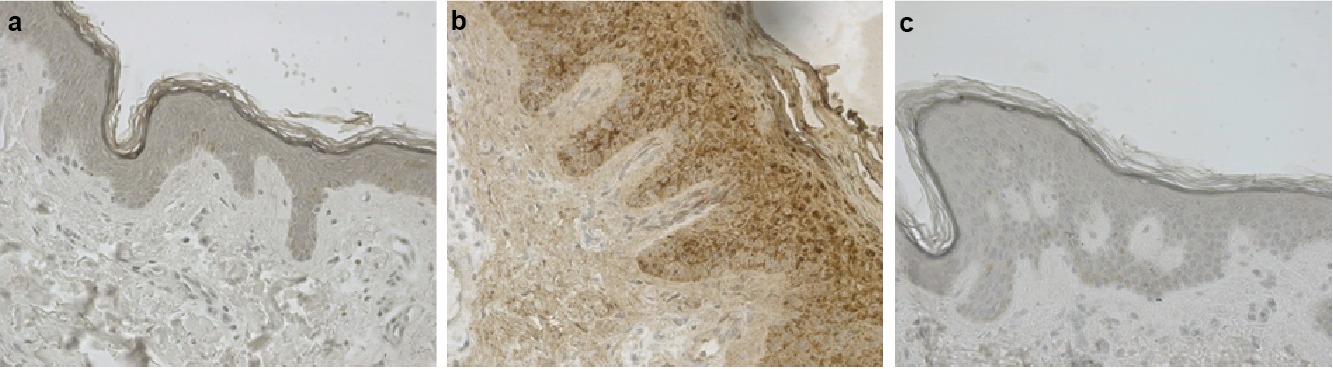 Click to show fullsize
Click to show fullsize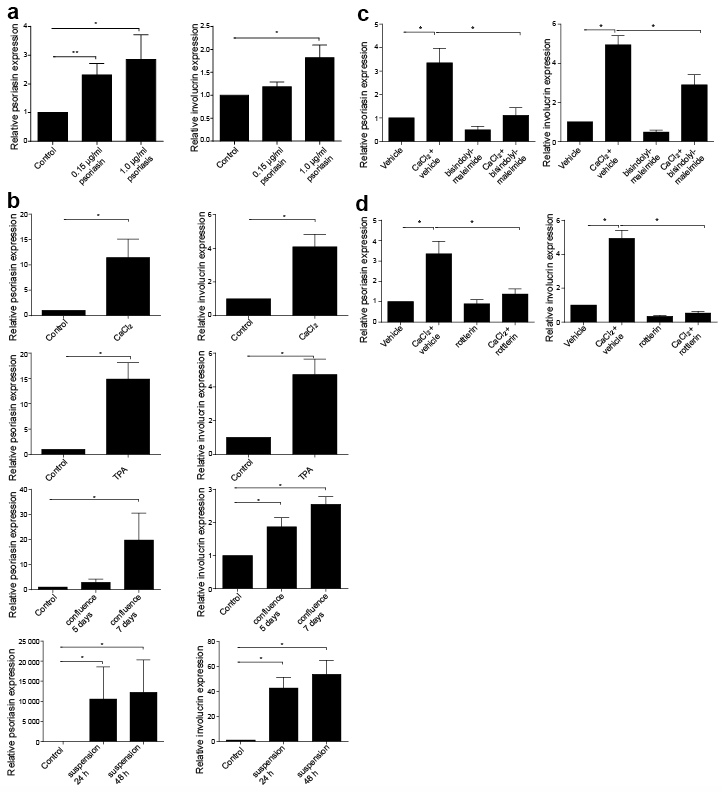 Click to show fullsize
Click to show fullsize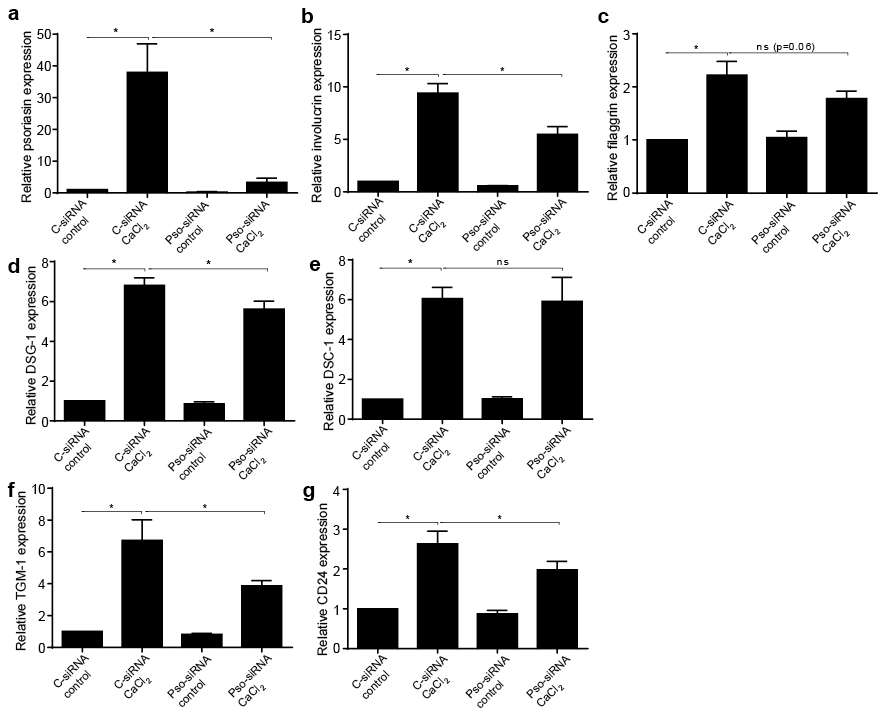 Click to show fullsize
Click to show fullsize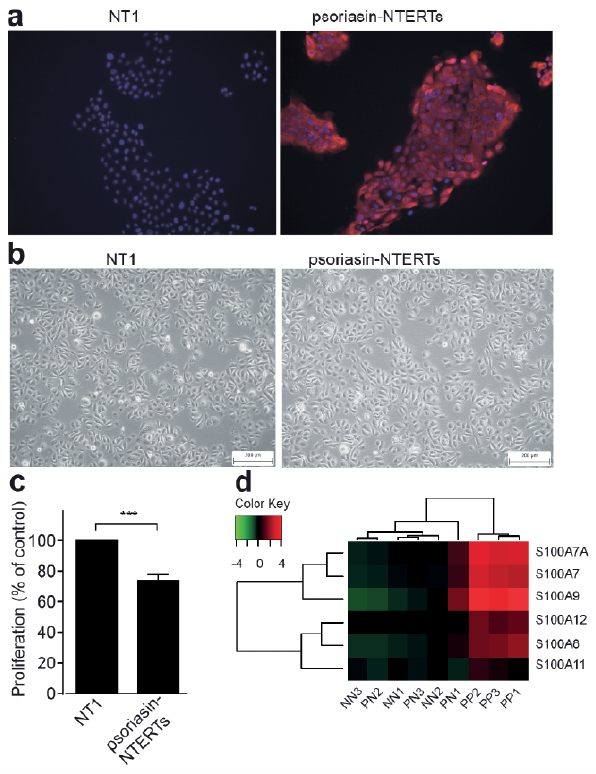 Click to show fullsize
Click to show fullsize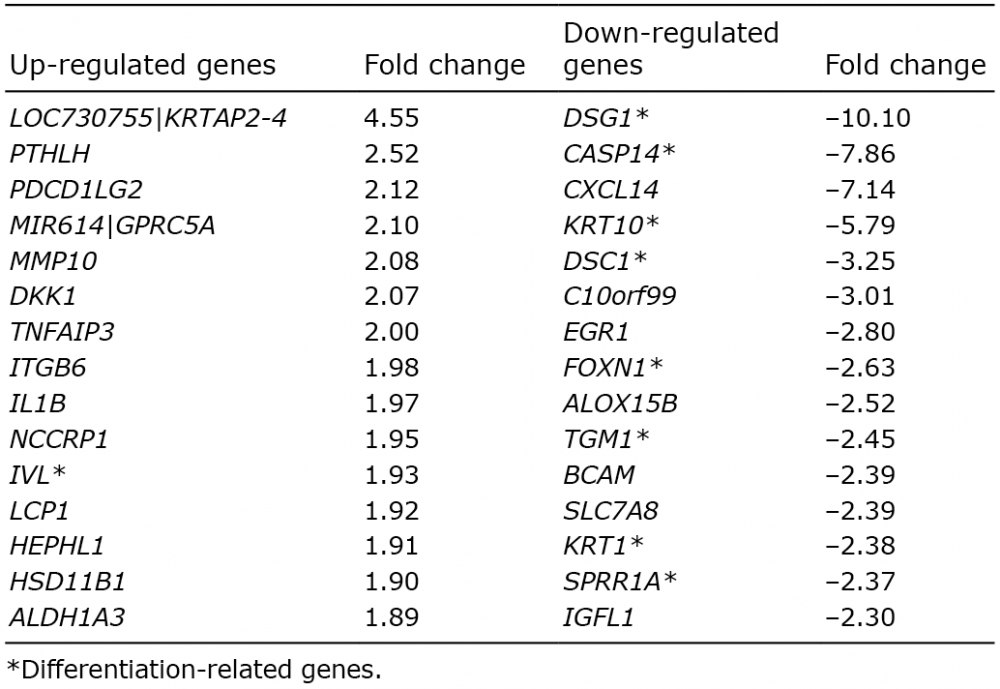 Click to show fullsize
Click to show fullsize