


Ingrid Asp Psoriasis Research Center, Department of Clinical and Experimental Medicine, Faculty of Medicine and Health Sciences, Linköping University, Linköping, Sweden
Psoriasis is a common autoimmune skin disease. The aim of this study was to investigate whether the apop-totic process is disturbed in psoriatic keratinocytes. In vitro culture of keratinocytes derived from both involved and uninvolved psoriatic skin, revealed higher viability and resistance to apoptosis following exposure to ultraviolet B, compared with cells from healthy controls. The position of apoptotic dysregulation was found to be upstream of cytochrome c release in the mitochondrial apoptotic pathway. Microarray transcriptome analysis revealed that 87 genes were differentially expressed in both involved and uninvolved psoriatic keratinocytes compared with controls. Among these, a general upregulation of anti-apoptotic genes and downregulation of pro-apoptotic genes were identified. This distinct apoptosis-resistant phenotype, unrelated to the inflammatory component of the disease, implies that intrinsic abnormalities in keratinocytes may contribute to the pathogenesis of psoriasis.
Key words: psoriasis; keratinocytes; apoptosis.
Accepted Mar 22, 2017; Epub ahead of print Mar 28, 2017
Acta Derm Venereol 2017; 97: .
Corr: Charlotta Enerbäck, Ingrid Asp Psoriasis Research Center, Department of Clinical and Experimental Medicine, Faculty of Medicine and Health Sciences, Linköping University, SE-581 85 Linköping, Sweden. E-mail: charlotta.enerback@liu.se
Psoriasis is a chronic inflammatory skin disease, affecting 2–3% of the population worldwide (1). Plaque psoriasis develops into red scaly patches, characterized by a dense inflammatory infiltrate, hyperproliferation of the epidermal keratinocytes, and increased angiogenesis. Overexpression of psoriasin (S100A7) contributes to dysregulated differentiation in psoriasis (2). The current general paradigm defines psoriasis as a T-cell-mediated disease targeting keratinocytes. However, there is evidence that identifies keratinocytes as important contributors to the pathogenesis of psoriasis.
A number of studies report different keratinocyte properties, not only in involved psoriatic skin, but also in normal-appearing uninvolved psoriatic skin that shows no signs of inflammation compared with normal skin. This suggests that an intrinsic abnormality in the keratinocytes contributes to the pathogenesis of the disease. Accelerated wound healing (3) and a disturbed calcium metabolism (4) have been demonstrated in uninvolved psoriatic skin compared with normal skin, as well as a hyper-responsiveness of uninvolved keratinocytes to growth-promoting T-cell lymphokines (5). A global gene expression profiling study demonstrated a “pre-psoriatic” gene expression signature in normal-appearing psoriatic skin (6), while another array study identified 562 genes that were differentially expressed in normal vs. uninvolved skin (7).
Programmed cell death, or apoptosis, is an essential process to eliminate damaged, dysfunctional or excessive cells. It is a multistep process regulated by a large number of intracellular and extracellular events, triggered by a variety of stimuli, such as DNA damage, the loss of adhesion, and growth factor withdrawal (8). A reduced apoptotic index has been reported in psoriatic epidermis compared with normal skin (9). Apoptosis resistance in psoriatic epidermal cells has previously been suggested by Wrone-Smith et al. (10), who showed that cells from psoriatic plaques had a prolonged capacity to resist apoptosis induced by methyl-cellulose suspension. It has since been accepted that apoptosis resistance may exist in psoriatic keratinocytes, although this assumption is based on very limited data.
In order to characterize intrinsic abnormalities in keratinocytes that contribute to the pathogenesis of psoriasis, this study investigated the apoptotic capacity of psoriatic keratinocytes. When comparing the apoptotic stress response induced by ultraviolet B (UVB) irradiation between psoriatic and normal keratinocytes, a significantly reduced amount of apoptosis was found in the cells derived from psoriatic skin, including both involved and uninvolved areas. Microarray analysis further demonstrated preferential upregulation of anti-apoptotic genes, as well as downregulation of pro-apoptotic genes, which could reflect the observed apoptosis resistance.
All patients and control subjects included in the study were examined and the diagnosis of psoriasis verified by a dermatologist at the Department of Dermatology at Linkoping University Hospital. The study was approved by the local ethics committee. All participants gave their written informed consent.
In order to perform experiments on living psoriasis keratinocytes, we have established a cell culture system, as described previously, with some modifications (11). Tissue samples were obtained as 4-mm punch biopsies of the skin from both involved and uninvolved skin areas (matched samples) of patients with psoriasis. In addition, biopsies were taken from healthy control individuals. The biopsies were taken from the non-UV-exposed buttock region. In all, the experiments included primary cultures of keratinocytes from 10 patients and 10 healthy controls. The number (n) in the figure legends indicates the number of experiments that have been performed, with each n being keratinocytes established from a separate individual.
After incubation with dispase (2 mg/ml) at 4°C for 18 h or with ammonium thiocyanate (3.8%) at room temperature for 30 min, the epidermal layer was separated from the dermal part of the skin and placed in 0.05% trypsin for 40 min. The cells were then filtered through a 40 mm nylon cell strainer, pelleted and resuspended in cell culture medium. The keratinocytes were cultured in Dulbecco’s modified Eagle’s medium (DMEM)-Ham’s F-12 (3:1) with 10% foetal bovine serum supplemented with penicillin (50 U/ml), streptomycin (50 µg/ml) and fungizone (50 µg/ml) (all from Invitrogen, Paisley, UK), insulin (5 µg/ml), cholera toxin (0.1 nM), hydrocortisone (0.4 µg/ml), triiodothyronine (2 nM), transferrin (5 µg/ml) and adenine (0.18 mM) (all from Sigma-Aldrich, St Louis, MO, USA), and epidermal growth factor (10 ng/ml) (Roche Diagnostics, Mannheim, Germany), according to Rheinwald (12). Differential trypsinization was performed to obtain pure cultures. The cells were grown in monolayers in Falcon tissue culture dishes (BD Biosciences, San Jose, CA, USA) and maintained at 37°C in a humidified atmosphere of 5% CO2 in air. Pure keratinocyte cultures were established after 2–3 passages and the experiments were conducted at passage 4. The experiments were performed with a seeding density of 15,000 cells/cm2. Untreated controls from the same individual were analysed in parallel.
Apoptosis was induced by UV radiation, a well-described model for apoptosis induction in keratinocytes in vitro (13). The UVB source comprised 2 Philips TL20W/12 tubes (Philips, Eindhoven, the Netherlands) emitting in the 280–370 nm range, with a main output of 305–320 nm. A Schott WG 305 cut-off filter (50% absorption below 305 nm, Mainz, Germany) was used. The output was 1.44 mW/cm2, measured with a psoralen plus ultraviolet A (PUVA) Combi Light dosimeter (Leuven, Belgium), using adequate adjustments for UVB. The irradiation was performed in phosphate-buffered saline (PBS) and no temperature increase in the PBS was observed during irradiation. Directly after irradiation, the PBS was replaced by fresh pre-warmed (37°C) medium. The keratinocytes were irradiated 18 h after seeding with the UV doses of 200 and 400 mJ/cm2 and collected 6 h and 24 h post-irradiation, respectively. Non-irradiated control cells were handled identically, but not irradiated.
Cell viability was measured using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) reduction assay (Calbiochem, San Diego, CA, USA). Cells were incubated with 0.25 mg/ml MTT for 2 h at 37°C. Absorbance was measured at 550 nm.
To analyse caspase-3 activity, the keratinocytes were permeabilized in lysis buffer (10 mM Tris-HCl pH 7.5, 1% Triton X-100, 130 mM NaCl, 10 mM NaH2PO4/NaHPO4, 10 mM sodium pyrophosphate) 6 h after UV irradiation and incubated with the Ac-DEVD-AMC substrate, according to the manufacturer’s recommendation (BD Pharmingen, San Diego, CA, USA). The concentration of proteolytically released AMC substrate (7-amino-4-methylcoumarin) was analysed in a Shimadzu RF-540 spectrofluorometer (λex380/λem435, Shimadzu, Kyoto, Japan). The fluorescence was correlated with protein content.
Cytosol was extracted from the keratinocytes by adding digitonin (Sigma-Aldrich) diluted in a buffer consisting of 250 mM sucrose, 20 mM Hepes, 10 mM KCl, 1 mM EDTA, 1 mM EGTA, 1.5 mM MgCl2, 1 mM Pefabloc and 8 mM dithiotriol, pH 7.5. This treatment permeabilizes the plasma membrane but leaves intracellular organelle membranes intact, due to the proportionally higher cholesterol content of the plasma membrane. This was verified by the analysis of lactate dehydrogenase (LDH) and the lysosomal enzyme β-N-acetylglucoseaminidase (NAG) activity (14, 15). The digitonin concentration (10–15 µg/ml) was individually titrated for each keratinocyte donor. Protein extraction was performed by trichloric acid (50%) precipitation. The pellet was then resuspended in urea-lysis buffer (6 M urea, 150 mM NaCl, 5 mM EDTA, 0.1% SDS, 1% Triton X-100, 50 mM Tris, pH 8.0) and 1 M NaOH).
Samples containing 20 μg protein lysate were loaded onto mini-PROTEAN TGX precast gels (Bio-Rad Laboratories, CA, USA) and transferred to polyvinylidene difluoride membranes in a Trans-Blot Turbo system (Bio-Rad Laboratories). Subsequently, the blots were saturated with 5% BSA and primary monoclonal mouse cytochrome c (1:400, BD Pharmingen) and p53 (1:1000, Santa Cruz Biotechnology, Dallas, TX, USA) antibodies or polyclonal rabbit phos-p53 (Ser15, 1:1000, Cell Signaling, Danvers, MA, USA) antibodies were used, followed by horseradish peroxidase (HRP)-conjugated sheep anti-mouse or donkey anti-rabbit secondary antibodies (1:5000 (1:2000 cyt c), Amersham Biosciences, Buckinghamshire, UK). The membranes were developed with the enhanced ECL-Plus Western blotting detection system (Amersham Biosciences) and were re-probed with mouse glyceraldehyde-3-phosphate dehydrogenase (GAPDH; 1:1000, Biogenesis, Poole, UK) as an internal control. ImageJ software (National Institutes of Health, Bethesda, MD) was used to measure the optical density. All the results were normalized to the amount of GAPDH.
Keratinocytes were re-suspended in RLT Lysis buffer (Qiagen, Valencia, CA, USA) containing 1% β-mercaptoethanol and the RNA was isolated using the RNeasy Plus Mini kit (Qiagen), according to the manufacturer’s instructions. Purified keratinocyte RNA (300 ng) was reverse transcribed to cDNA using the Maxima™ first-strand cDNA synthesis kit (Thermo Scientific Fermentas, Vilnius, Lithuania). The reverse transcriptase reaction was performed in a total volume of 20 µl at 25°C for 10 min, 50°C for 15 min and 85°C for 5 min.
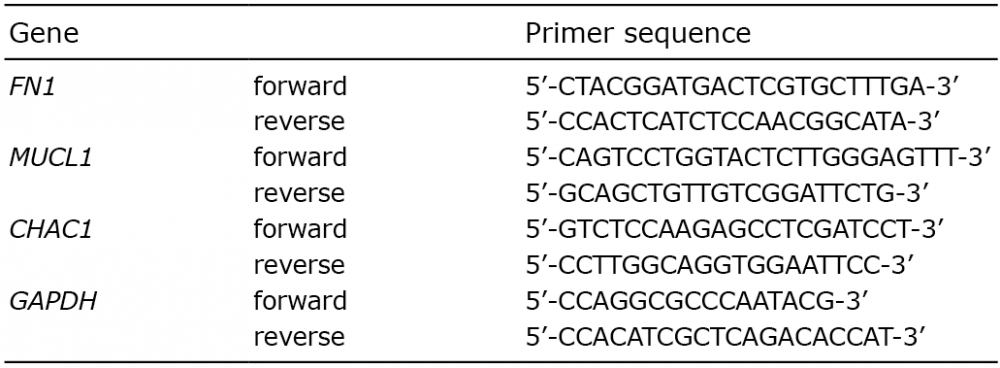
Gene expression of FN1, MUCL1 and CHAC1 were quantified with the Power SYBR® Green PCR Master Mix (Applied Biosystems, Foster City, CA, USA) in the ABI 7500 Fast Real-Time PCR System (Applied Biosystems). The sequences for the primers that were used are listed in Table I (Life Technologies, Bleiswijk, The Netherlands). Amplifications were performed in a total volume of 20 μl for 1 cycle of 50°C for 2 min, 1 cycle of 95°C for 10 min, and 40 cycles of 95°C for 15 s and 60°C for 1 min. Samples were run in triplicate and the relative mRNA expression of the gene of interest relative to the housekeeping gene, GAPDH, for each sample was expressed as 2–?Ct.
Table I. Primer sequences
The Affymetrix GeneChip Human Transcriptome Array 2.0 (HTA 2.0) is a high-resolution array, including more than 6 million probes targeting coding transcripts, non-coding transcripts and exon-exon splice junctions. A total of 250 ng of RNA were processed using the GeneChip® WT Plus Reagent Kit (Affymetrix, Santa Clara, CA, USA), according to the Affymetrix protocol. The Affymetrix HTA 2.0 microarrays were hybridized in a GeneChip® hybridization oven at 45°C, at 60 rpm, for 16 h. The washing and staining procedure was performed in an Affymetrix GeneChip® Fluidics Station 450 and the Affymetrix GeneChip® Scanner 3000 7G system was used to scan the arrays. Internal controls of the hybridized chips were verified and the CEL files generated by the Affymetrix AGCC program were evaluated using GeneSpring GX software (version 13.0; Agilent Technologies, Inc., Santa Clara, CA, USA). Standard Robust Multi-array Average (RMA) normalization and background correction were performed and the data log was transformed. The data have been deposited in the NCBI Gene Expression Omnibus (16) and are accessible through GEO Series accession number GSE93423. Differentially expressed genes were identified through fold change using the threshold of ≥ 1.5. To explore the functions of the differentially expressed genes, gene ontology (GO) trees were constructed with regard to the biological processes, molecular functions and cellular components of the proteins encoded by the genes. Specific gene groups were then extracted and evaluated further.
Data analysis was performed with GraphPad Prism 6 (GraphPad, San Diego, CA, USA). Statistical comparisons were performed using a Student’s t test for unpaired and paired samples. p-values below 0.05 were considered significant.
First, we aimed to investigate the survival response of UVB-irradiated psoriatic and healthy control keratinocytes. After 6 h, we demonstrated a significantly higher viability rate for UVB-exposed (200 mJ/cm2) keratinocytes established from involved and uninvolved psoriatic skin compared with healthy control skin (p = 0.030 and p = 0.016 respectively) (Fig. 1a). In addition, the keratinocytes from uninvolved skin displayed a higher viability after UVB irradiation, compared with control cells. The same trend was shown in cells harvested after 24 h (data not shown). Following exposure to the higher UV dose of 400 mJ/cm2, the response pattern was similar, with borderline significance (p = 0.067), in the comparison of viability between involved psoriatic keratinocytes and control keratinocytes. The mean ± standard error of the mean (SEM) absorbance value of the control cells was 0.416 ± 0.078.
Fig. 1. Apoptotic dysregulation in psoriatic keratinocytes (KC) in response to ultraviolet B (UVB) irradiation. (a) Psoriatic (involved and uninvolved) KC and healthy KC were exposed to UVB radiation (200 mJ/cm2, 400 mJ/cm2). Viability was analysed with an 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) viability assay 6 h after UVB exposure and it is presented as the percentage of viability compared with unexposed cells. *p ≤ 0.05, **p ≤ 0.01, n = 6–7, mean ± standard error of the mean (SEM). (b) Psoriatic (involved and uninvolved) KC and healthy KC were exposed to UVB radiation. Caspase-3 activity was detected as the cleavage of the Ac-DEVD-AMC substrate 6 h after UVB exposure. *p ≤ 0.05, n = 7–9, mean ± SEM. (c) Western blot analysis of cytochrome c in cytosolic fractions of psoriatic (involved and uninvolved) KC and healthy KC 6 h after 200 mJ/cm2 UVB radiation. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as an internal control and one representative blot is shown. A densiometric analysis of cytochrome c correlated to the GAPDH level is presented as the fold increase compared with unexposed control cells. n = 2, mean ± SEM.
To determine whether the observed UV resistance was due to inhibited apoptosis, a caspase-3-activity assay with the caspase-3-specific substrate, Ac-DEVD-AMC, was performed. UVB exposure induced caspase-3 activation, which indicates that apoptotic cell death takes place. The results presented in Fig. 1b clearly demonstrate that the apoptotic response between the samples differs. Cells from healthy control skin displayed distinct caspase-3 activity 6 h following UVB exposure (200 mJ/cm2), while the activity in keratinocytes from both involved and uninvolved psoriatic skin was much lower compared with controls (p = 0.014 and p = 0.021 respectively). In addition, there was a significant difference in activity between involved and uninvolved skin keratinocytes (p = 0.040). The same trend was shown in cells harvested after 24 h (data not shown). The observed response pattern between psoriatic keratinocytes and controls was also present following the higher UVB exposure of 400 mJ/cm2. However, only borderline statistical significance was reached due to variations between the controls (p = 0.072 (involved), p = 0.087 (uninvolved)). The caspase-3 activity was 0.400 arbitrary units/mg protein in untreated control cells. There was no difference in viability or caspase-3 activity in non-irradiated cells from psoriatic and healthy skin, respectively (data not shown).
In order to localize the position at which the differential apoptotic response originates, cytosolic fractions were isolated from the cells using digitonin. Digitonin permeabilizes the plasma membrane, but leaves intracellular organelle membranes intact. UVB irradiation markedly increased the cytosolic cytochrome c protein level in keratinocytes from healthy control skin, as identified by Western blotting (Fig. 1c). Psoriatic keratinocytes, from both involved and uninvolved skin, displayed markedly less cytochrome c protein release from the mitochondria compared with healthy control cells. Our data demonstrate that the dysregulation in the apoptotic response is situated upstream of the mitochondrial cytochrome c release within the intrinsic apoptotic pathway.
To explore the transcriptional pattern that underlies the differences in the apoptotic response, we used the Affymetrix GeneChip Human Transcriptome Array 2.0 (HTA 2.0) to compare mRNA expression in UVB- and non-UVB-treated psoriatic and normal keratinocytes. The hierarchical clustering of gene expression in control and psoriatic keratinocytes revealed that the largest difference in gene expression was between UVB- and non-UVB-exposed cells rather than between patients and controls (Fig. 2a). In the presence of UV radiation, the samples cluster according to whether the skin is involved or uninvolved, rather than the individual from whom they were derived.
Fig. 2. Microarray analysis of ultraviolet B (UVB)-exposed psoriatic keratinocytes (KC). (a) Hierarchical clustering of gene expression in healthy control and UVB (200 mJ/cm2)-/non-UVB-exposed psoriatic KC. The Venn diagrams present the number of differentially expressed genes in (b) UVB-exposed and (c) non-UVB-exposed psoriatic KC from involved and uninvolved skin areas compared with healthy control KC. (d) The Venn diagram shows differentially expressed genes in involved psoriatic KC compared with uninvolved KC in non-UVB-exposed cells and UVB-exposed cells, respectively. Apoptosis-related genes that are differentially expressed in (e) UVB-exposed (200 mJ/cm2) psoriatic KC and (f) non-irradiated psoriatic KC compared with healthy control KC. The threshold for differentially expressed genes was a fold change ≥ 1.5, involved psoriatic KC (solid bars), uninvolved psoriatic KC (diagonally striped bars), n = 2. (g) Quantitative real-time reverse transcription PCR (qRT-PCR) analysis of selected transcripts (CHAC1, FN1 and MUCL1) in UVB-exposed psoriatic and healthy control KC. p ≤ 0.05, n = 4, mean ± standard error of the mean (SEM).
The Venn diagram demonstrates that 515 and 176 genes were differentially expressed in UVB-irradiated involved and UVB-irradiated uninvolved psoriatic keratinocytes, respectively, compared with UVB-exposed healthy control cells (Fig. 2b). Eighty-seven of these genes overlapped between involved and uninvolved samples. The corresponding numbers for non-UVB-exposed cells were 579 genes for involved and 155 genes for uninvolved psoriatic keratinocytes, and 78 of these genes overlapped (Fig. 2c). When comparing involved psoriatic keratinocytes with uninvolved keratinocytes, 287 and 260 genes were differentially expressed in non-UVB-exposed cells and UVB-exposed cells, respectively (Fig. 2d). A total of 160 of these genes overlapped. Inter-individual variations were approximately 1%.
We sought to identify genes differentially expressed in both involved and uninvolved cells, since we hypothesized that the same mechanism was responsible for the apparent apoptosis resistance in both cell types. Of the 8 most upregulated genes in involved psoriatic skin, 4 were among the 20 most upregulated genes in uninvolved skin (Table II). These genes were MUCL1 (mucin-like 1), FN1 (fibronectin), UPK1B (uroplakin 1B) and CXCL5 (chemokine (C-X-C motif) ligand 5). Similarly, among the genes downregulated in both UVB-irradiated psoriasis-involved and -uninvolved keratinocytes, compared with normal keratinocytes, we identified CHAC1 (ChaC glutathione-specific gamma-glutamylcyclotransferase 1) and GSDMC (gasdermin C).
Table II. The most up- and downregulated genes in ultraviolet (UV)-exposed involved and uninvolved psoriatic keratinocytes vs. UV-exposed healthy control keratinocytes
Next, we selected genes defined as being apoptosis related by GeneSpring software. Figs 2e and 2f show the expression of the apoptotic regulatory genes that were differentially expressed in psoriatic keratinocytes compared with healthy controls. Interestingly, the large majority of the anti-apoptotic genes were upregulated in the psoriatic cells in response to UVB radiation, while almost all the pro-apoptotic differentially expressed genes were downregulated in these cells. This imbalance between pro- and anti-apoptotic gene expression is in line with the observed apoptosis resistance in these cells. In contrast, this expression pattern was not observed in non-UVB-exposed keratinocytes. Here, the variable expression of anti- and pro-apoptotic genes probably represents the homeostasis between cell death and survival present in non-stressed cells. In addition, the Bax/Bcl2 ratio was extracted from the microarray data. The ratio was close to 1, both when comparing non-UV and UV exposed psoriatic and control cells. Thus, the observed apoptosis resistance cannot be explained by a skewed Bax/Bcl2 ratio.
To confirm and validate the microarray results, we performed quantitative real-time reverse transcription PCR for the FN1, MUCL1 and CHAC1 genes in UVB-exposed psoriatic and healthy control keratinocytes (Fig. 2g). CHAC1 and FN1 were upregulated 2.1-fold (p = 0.048) and 2.4-fold (p = 0.027) respectively, in psoriatic keratinocytes relative to healthy control cells. MUCL1 was downregulated 3.5-fold without reaching statistical significance. These results uniformly confirmed the results of the microarray analysis.
p53 has been implicated in UV-induced apoptosis (17, 18). We therefore further explored the apoptotic role of p53 in UVB-exposed psoriatic keratinocytes, although p53 was not among the most differentially expressed genes in the microarray analysis. The protein expression of phosphorylated p53 (Ser15), implicated in apoptosis induction in response to UV radiation, was determined using Western blotting. UVB irradiation induced phosphorylation in both healthy and psoriatic cells, but no significant difference was observed between them (Fig. 3a). Next, the total protein expression of p53 was analysed. Following UVB irradiation, the total p53 protein level increased in control cells (p = 0.039, Fig. 3b).
Fig. 3. p53 expression in psoriatic keratinocytes following ultraviolet B (UVB) irradiation. (a) Western blot analysis of (a) phosphorylated p53 (Ser15) and (b) total p53 protein expression 6 h after UVB exposure (200 mJ/cm2) in involved and uninvolved psoriatic keratinocytes and healthy control keratinocytes (KC). A densitometric analysis of phosphorylated p53 and total p53 respectively correlated to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) level is presented as the fold increase compared with unexposed control cells. *p ≤ 0.05, **p ≤ 0.01, n = 4–6, mean ± standard error of the mean (SEM).
The same trend was observed for the psoriatic keratinocytes, but only uninvolved keratinocytes reached statistical significance (p = 0.017). The basal levels of p53 were significantly lower in psoriatic keratinocytes compared with cells from healthy skin (p = 0.006 uninvolved keratinocytes, p = 0.034 involved keratinocytes), but the biological significance of this observation is unclear. The p53 results at mRNA, protein and phosphorylation levels exclude p53 as an important factor in the resistance to apoptosis observed in psoriatic keratinocytes.
To analyse the expression of apoptotic genes in psoriatic skin, we performed a microarray transcriptome analysis of the epidermis from psoriatic patients and control individuals (n = 3). Differentially expressed genes (≥ 1.5-fold) were selected for Gene Ontology terms associated with apoptosis or the regulation of apoptosis. A number of apoptosis-related genes were found to be differentially expressed, which further supports the concept of a dysregulated apoptotic process in psoriasis (Table III).
Table III. Differentially expressed apoptotic genes in psoriatic and control epidermis
The gene that demonstrated the strongest differential expression (upregulated 17.8 times) was IFI6 (interferon alpha-inducible protein 6) and the second most strongly expressed gene, upregulated 12.1 times was IFI27 (interferon alpha-inducible protein 27). Pathway analysis using GeneSpring did not result in the identification of any pathway clusters, probably due to small sample size.
This study used a cell culture system that selects for the proliferative compartment of psoriatic keratinocytes as an in vitro model to study psoriasis. It was demonstrated that, in response to the physiological stress agent, UVB, both involved and uninvolved psoriatic keratinocytes respond with a lower level of apoptosis compared with keratinocytes from normal, healthy skin. These differences in apoptotic susceptibility do not depend primarily on exogenous influences but instead on an intrinsic cellular defect, since the observed alterations remained after several passages in culture. The presence of intrinsic abnormalities in psoriatic keratinocytes are supported by genetic association studies showing strong association of genes specifically expressed in keratinocytes, such as late cornified envelope (LCE) and CARD14 (19, 20). However, a persisting effect of infiltrating T-cells may not be ruled out.
Wrone-Smith et al. (10) cultured psoriatic and normal keratinocytes in methyl cellulose to study the effect of contact inhibition. In contrast to normal skin and skin from uninvolved areas, they found no double-stranded DNA fragmentation in keratinocytes derived from psoriatic plaques. It was suggested that the psoriatic plaque keratinocytes had a prolonged capacity to resist apoptosis compared with normal keratinocytes. The fact that keratinocytes from psoriatic skin in this and our study demonstrate a similar response when exposed to 2 very distinct forms of stress stimulus implies that these cells have a genuine apoptosis-resistant nature.
Several factors, such as UV dose, balance between death and survival factors and variations in intracellular signalling, determine whether or not keratinocytes will undergo UV-induced apoptosis. An apoptosis-inducing dose of UV radiation may be counteracted by an increased amount of survival-promoting molecules. The microarray data reflect that the majority of the differentially expressed genes belonging to the anti-apoptotic family were expressed to a higher degree after UV treatment in psoriatic keratinocytes compared with controls. Likewise, the majority of the pro-apoptotic genes were expressed to a lower degree in the keratinocytes from psoriatic skin. Together, these events clearly shift the balance towards a more survival-promoting capacity.
We show that UVB radiation induce the mitochondrial pathway of apoptosis in keratinocytes, as detected by cytochrome c release from the mitochondria to the cytosol and caspase-3 activity, which is in agreement with previous findings (13, 17). Our findings further suggest that the cause of the deregulated apoptosis in psoriatic keratinocytes is located upstream of cytochrome c release. There are very few studies addressing the underlying mechanisms of the proposed cell death resistance of psoriatic keratinocytes. Reports on the expression of apoptosis-related proteins in psoriasis are divergent. Several immunohistochemical studies demonstrate a strong Bcl-X expression in both involved and uninvolved psoriatic skin, significantly higher than in the epidermis of healthy skin (21, 22). Bcl-2, another anti-apoptotic protein belonging to the same protein family, has also been reported to be upregulated in psoriasis (23). How-ever, contradictory results have shown that Bcl-2 is reduced in involved psoriatic skin (21, 22). The overexpression of the pro-apoptotic Bax and Bak proteins has also been found in psoriatic plaques (21, 22, 24) and it has been suggested to reflect an inhibitory mechanism counteracting the intensive proliferation (21).
In the microarray of cultured keratinocytes, MUCL1 was the most upregulated gene in both involved and uninvolved psoriatic cells. It has been suggested that MUCL1 is a biomarker of diagnosis in metastatic breast cancer (25), where its expression correlated to the psoriasis-associated protein, psoriasin (S100A7) (26). FN1 (fibronectin) was also one of the most upregulated genes in both involved and uninvolved skin. FN1 has anti-apoptotic properties and stimulates cell proliferation through the induction of AKT phosphorylation and the increased DNA binding of nuclear factor (NF)-?B (27). The extra domain A (EDA) isoform of FN (EDA+ FN), as well as molecules involved in FN1-mediated integrin signalling, have previously been shown to be overexpressed in psoriasis, in both involved and uninvolved epidermis (28, 29). The strong overexpression suggests a role in the observed anti-apoptotic properties of psoriatic keratinocytes. Another gene upregulated in both involved and uninvolved skin was CXCL5, which is a chemokine involved in the recruitment and activation of leukocytes during inflammation. CXCL5 can be induced in keratinocytes by IL17A and has been shown to be highly expressed in psoriatic keratinocytes (30). Among the 90 different inflammatory mediators measured in a study by Dawes et al. (31), CXCL5 was shown to be induced to the greatest extent by UVB treatment in human skin. Interestingly, we found an increase in CXCL5 expression after UVB treatment, which was not present in cultured keratinocytes before UV. CHAC1 was downregulated in both involved and uninvolved psoriatic cells compared with controls and it is a recently identified member of the unfolded protein response (UPR) pathway, which is a stress-signalling pathway in the endoplasmic reticulum (32). It has been characterized as a pro-apoptotic component of this pathway. The downregulation of the pro-apoptotic CHAC1 in psoriatic keratinocytes may contribute to the apoptosis resistance in these cells. Madonna et al. (33) have demonstrated that the suppressor of cytokine signalling (SOC1 and -3) suppresses cytokine-induced apoptosis in psoriatic keratinocytes, but these molecules were not differentially expressed in our experimental setting.
P53 has been shown to be an important regulator of UVB-induced apoptosis in keratinocytes (18). In mice deficient in p53, the number of observed apoptotic keratinocytes, commonly named “sunburn cells”(34), in the skin was markedly reduced following UVB exposure (35). Stimuli, such as UV-induced DNA damage, lead to the multisite phosphorylation of p53, primarily through the phosphorylation of serine 15 (36). This phosphorylation site has been shown to be implicated in the induction of apoptosis by p53 (37). In contrast to the suggested importance of p53 in UVB-induced apoptosis, our results did not indicate that p53 plays a central role in the observed apoptotic resistance.
We performed a microarray transcriptome analysis of the epidermis from patients with psoriasis and control individuals to investigate the expression of apoptosis-regulated genes in the tissue. A number of these genes were found to be differentially expressed in psoriatic epidermis compared with controls, which further supports a dysregulation of the apoptotic process in psoriasis. IFI6, which demonstrated the strongest differential expression, upregulated 17.8 times, has previously been shown to be significantly increased in psoriasis (38). IFI6 inhibits apoptosis via the mitochondrial-dependent pathway (39) and may contribute to the observed apoptotic resistance in the psoriatic skin. The second most strongly expressed gene, IFI27, has also been reported to be highly expressed in psoriatic lesions (40, 41). IFI27 has been suggested to be pro-apoptotic in some cell types (42), but a recent study suggested IFI27 to be required for cell proliferation in epidermal keratinocytes, with no effect on apoptosis or on apoptosis-related genes in this cell type (43).
In conclusion, our data suggest that psoriatic keratinocytes possess an apoptotic-resistant ability following UVB irradiation compared with cells from healthy skin. Interestingly, keratinocytes established from normal-appearing uninvolved psoriatic skin also demonstrate UV resistance. These observed differences suggest that endogenous defects in keratinocytes may be involved in the pathogenesis of the disease. This remarkable feature, which was also observed in uninvolved keratinocytes, needs to be further elucidated in psoriasis.
This research was supported by the Swedish Research Council, the Ingrid Asp Foundation, the Welander Foundation and the Swedish Psoriasis Association.
The authors declare no conflicts of interest.


 Click to show fullsize
Click to show fullsize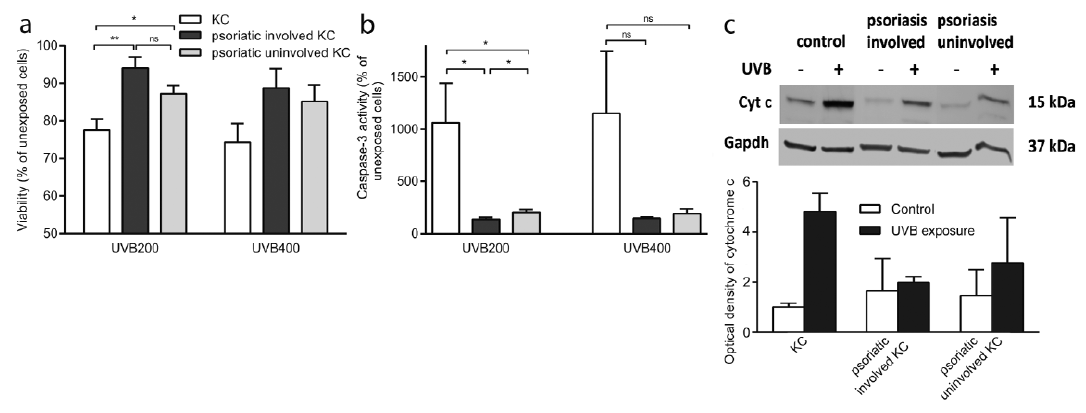 Click to show fullsize
Click to show fullsize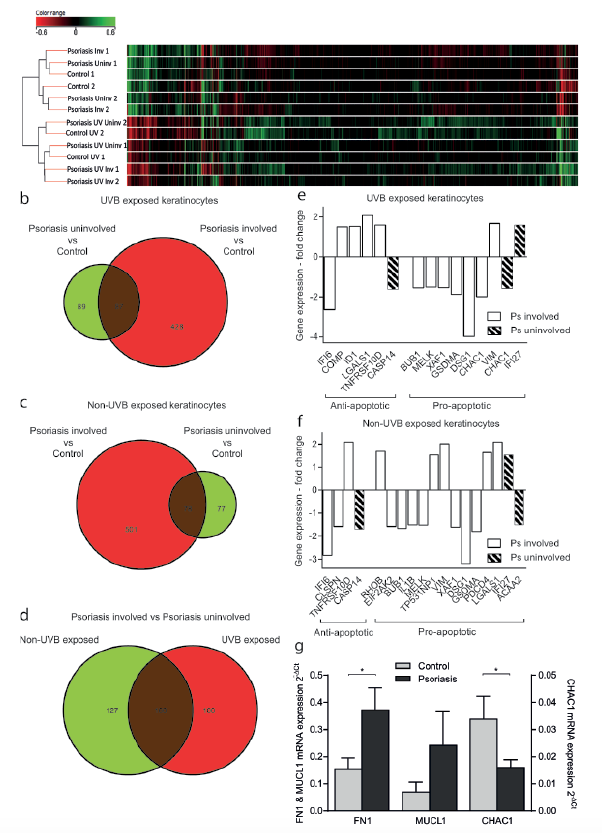 Click to show fullsize
Click to show fullsize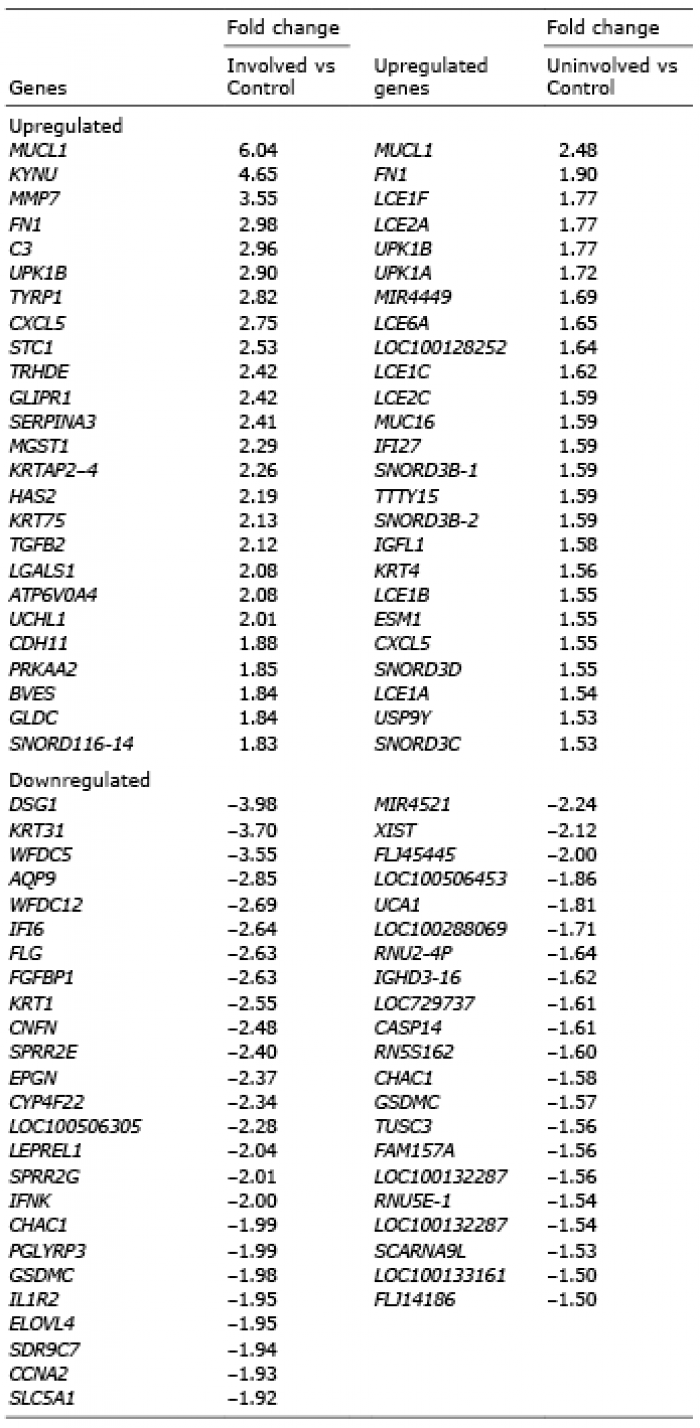 Click to show fullsize
Click to show fullsize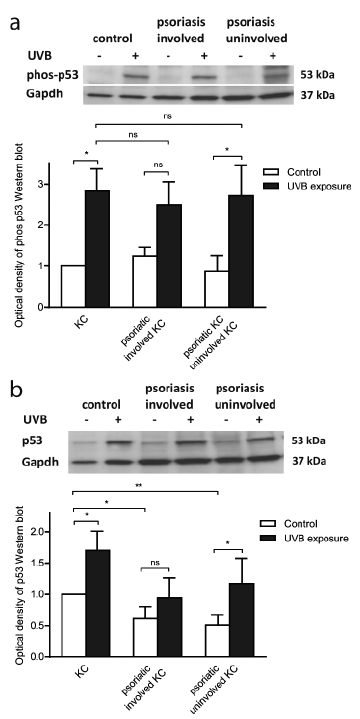 Click to show fullsize
Click to show fullsize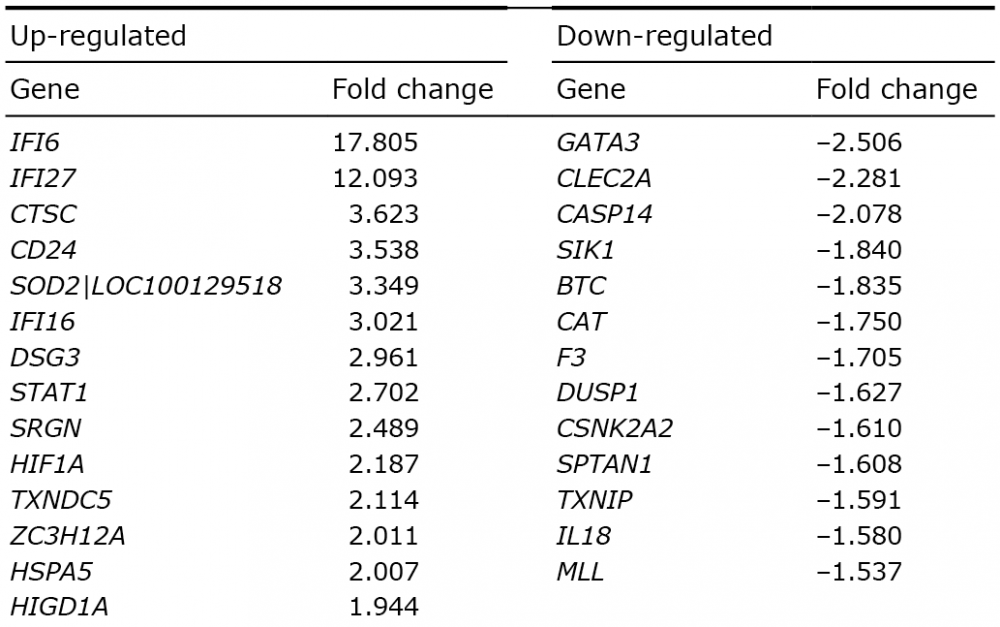 Click to show fullsize
Click to show fullsize