


Department of Molecular Biomedical Sciences, NC State Veterinary Medicine, 1060 William Moore Drive, Office 242, Raleigh, NC 27607, USA. *E-mail: skmishra@ncsu.edu. #These authors contributed equally.
Accepted May 22, 2018; Epub ahead of print May 24, 2018
Atopic dermatitis (AD) is one of the most common itch-inducing allergic skin diseases in humans and animals (1, 2). Interleukin (IL)-31, a cytokine involved in immune response in inflammatory diseases such as AD, has been demonstrated to cause itch by acting on receptors in a subset of transient receptor potential vanilloid (TRPV1) neurons (3–5). We recently identified a small subset of neurons that function as the primary detectors of chemical pruritic stimuli. We showed that the neuropeptide natriuretic polypeptide b (NPPB) is required for the transmission of itch signals and marks the full complement of itch responsive neurons at the periphery (6). However, it remains unknown how itch signals in AD are transmitted by the primary afferents of the sensory neurons in the dorsal root ganglia (DRG) to the spinal cord. This study is aimed to determine if NPPB is involved as a neuropeptide in IL-31-mediated itch in AD via natriuretic polypeptide receptor A (NPRA) in the spinal cord.
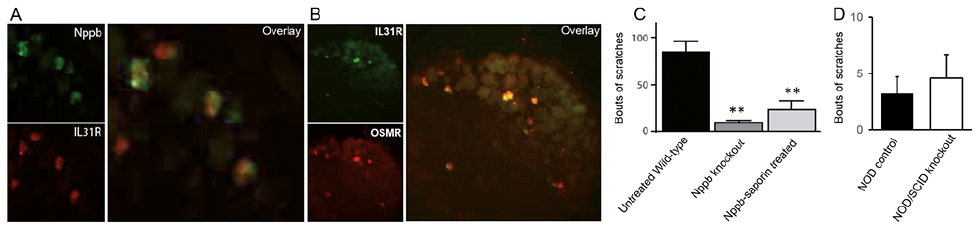
Cytokine IL-31 transmits its signals through a receptor complex made of IL31R and oncostatin M receptor (OSMR). We used double ISH to show that the IL31R is co-expressed with NPPB expressing sensory neurons in the DRG. Our data demonstrated that IL31R is selectively expressed in all NPPB-positive somatosensory neurons (Fig. 1A). We further showed that IL31R is exclusively expressed in all OSMR neurons in the DRG (Fig. 1B). We showed the co-expression of NPPB with IL31R in almost 98% of the neurons: 198/204 IL31R/OSMR-neurons.
Fig. 1. (A) Double in situ hybridization (ISH) reveals that the neuropeptide natriuretic polypeptide b (NPPB) (green) co-expresses interleukin (IL)-31-receptor (red). (B) Double ISH of IL-31Ra co-expresses oncostatin M receptor (OSMR). (C) Itch responses to subcutaneously injected IL-31 (1.5 nmol) were significantly attenuated in NPPB knockout mice and mice treated with NPPB-saporin. (D) No differences in itch responses were observed in mice lacking B- and T-cell receptors (NOD/SCID) compare to control mice (NOD). Itch behaviors were observed in mice within 30 min of administering IL-31. Each column represents the mean ± SEM, n ≥ 6 mice for each group, **p < 0.01.
To assess whether the IL-31-induced pruritogenic response is mediated via NPPB, we compared age-matched wild-type littermates and NPPB knockout (KO) mice. We injected IL-31 subcutaneously in both genotypes and measured scratching bouts. We observed a significant reduction in scratching response from NPPB KO mice (Fig. 1C). To further demonstrate the IL-31-mediated itch response by NPRA receptors expressed in the spinal cord, we used conjugated NPPB-saporin toxin to eliminate neurons expressing NPRA receptors in the spinal cord (6). We consistently found a drastic reduction in scratching response from the NPPB-saporin treated mice as compared to the control. To rule out immunogenic involvement of IL-31 in itch production, we administered IL-31 to NOD control mice and the mice lacking functional B and T-cells (NOD/SCID). We found no significant difference between NOD control (black bar) and NOD/SCID immune-deficient mice (white bar; Fig. 1D) suggesting IL-31-mediated itch is an immune-independent mechanism.
Contributions of TRPV1- or TRPA1-expressing DRG neurons in itch are important (8). Recently, individual TRPV1- or TRPA1-KO mice confirmed an important role of TRP-channels in IL31-induced itch (5); however, single KO have any compensatory effect on itch behavior was unclear. We sought out whether knocking out both TRPV1 and TRPA1-expression might show potential compensatory effect of TRP-channels on itch behavior. To determine the role of TRPV1/TRPA1 ion channels, we generated double KO mice. We first performed IL-31 induced calcium response on wild-type DRG sensory neurons (Fig. 2A). Quantification of the number of cells responding to IL-31 (0.3 μM) revealed a reduction (approximately 50%) in calcium response in double KO mice (Fig. 2B). Interestingly, our scratching behavior data demonstrated a significant decrease in IL-31 (0.3 nmol) induced itch response (80%) in TRPV1/TRPA1 double KO mice (Fig. 2C) compared to wild type mice. Next, we determined whether NPPB release is induced by cytokine IL-31 and if the release is mediated through TRPV1/TRPA1-ion channels. To explore this possibility, we cultured mouse DRGs from double KO and control mice and incubated neurons with IL-31 (0.3 μM) over 8 h to measure NPPB release. Indeed, our data showed a reduction (approximately 20%) in NPPB release in double KO mice as compared to control mice. These results demonstrate that both TRPV1 and TRPA1 channels are associated with NPPB-release in DRG sensory neurons.
Fig. 2. (A) Photomicrograph shows interleukin (IL)-31-evoked (0.3 μM) calcium signals in cultured sensory neurons following capsaicin (1 μM) as measured by Fura-2AM ratiometric calcium imaging. (B) Quantification of the percentage of IL-31-sensitive neurons in wild type and TRPV1/TRPA1 double knockout (KO) mice. Data show a significant reduction in calcium response (n = 3 mice) (total 450 neurons), *p <0.05. (C) Itch responses to subcutaneously injected IL-31 (0.3 nmol) in wild type, TRPV1/TRPA1 double KO mice (n ≥ 5 mice). Data show a significant reduction in scratching bouts between genotypes. **(unpaired Student t-test) were **p < 0.001. (D) Natriuretic polypeptide b (NPPB) release induced by IL-31 (0.3 μM) over 8 h was quantified by ELISA. A significant decrease in NPPB release in double KO mice compared to control was observed (n = 3 mice, *p < 0.05).
Acute itch sensation is mostly transient, caused by many factors, and generally relieved by anti-histamines. On the other hand, chronic itch is persistent and associated with many pathological conditions including allergic skin diseases (e.g., AD and psoriasis). In this study, we found a significant reduction in IL-31-induced scratching bouts from NPPB KO mice, suggesting a direct involvement of NPPB in IL-31-induced itch. This finding is corroborated by IL-31R co-expression in NPPB-positive neurons within the DRG. Our data indicate a simple molecular pathway and a highly specific cellular mechanism for itch in AD.
Recent studies have reported the co-localization of OSMR, a receptor subunit targeted by IL-31, in the non-peptidergic neuronal population (9). In contrast, along with other independent studies, we found immunoreactivity of IL-31 predominantly in peptidergic neuronal populations that overlapped with NPPB-positive neurons labeled by double ISH (Fig. 1) (5, 10).
Previously, it was shown that mast cells are not involved in IL-31-mediated itch response (5). However, it was not clear if B cells or T cells were involved in detecting skin itch. Our results did not observe a decrease in scratching bouts from NOD/SCID (B and T cells deficient mice) compared with NOD (control), indicating IL-31 could be acting through an immune-independent mechanism affecting the IL-31 receptors expressed on DRG neurons, as suggested by earlier studies (5, 10). A recent study showed that alone TRPV1- and TRPA1-positive neurons within the DRG co-express IL-31, but it remains unclear whether both TRPV1 and TRPA1 are essential in IL-31-mediated itch (5). Our study extends upon this previous finding by showing a significant reduction in IL-31-induced scratching bouts in TRPV1/TRPA1 double KO mice. We demonstrated a significant reduction in IL-31-induced calcium transients from DRG neurons in TRPV1/TRPA1 double KO mice, a finding supported by previous studies. Furthermore, our result reveals that IL-31 induced-NPPB release is mediated through TRPV1 and/or TRPA1-ion channels. It is worth noting that there was not a complete elimination of NPPB release to IL-31 and this could indicate a potential involvement of other ion-channels.
In conclusion, we have shown, for the first time, that neuropeptide NPPB is an important central component for itch transmission in allergic diseases like AD. Our results molecularly characterize the first two locations (NPPB in the DRG and NPRA-expressing neurons in the spinal cord) of an itch response pathway in mice induced through IL-31. We revealed an important role of neuropeptide NPPB in AD that could provide a therapeutic target for alleviating chronic itch associated with AD.
Funding supported by NC State Veterinary Medicine. NPPB KO mice were provided by Mark Hoon, NIDCR/NIH.
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize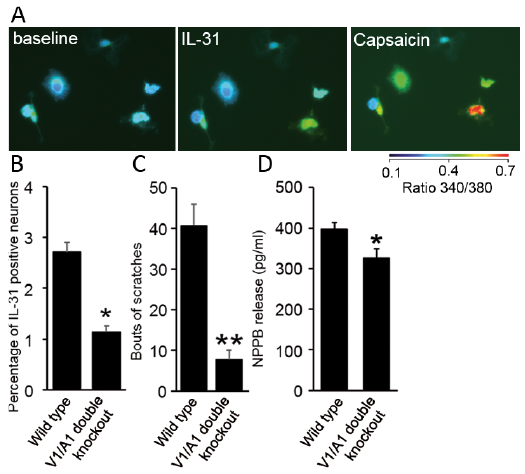 Click to show fullsize
Click to show fullsize