


1Department of Dermatology, Peking University First Hospital, Beijing, 2Department of Dermatology, Peking University Shenzhen Hospital, Shenzhen, Guangdong, and 3Shenzhen Key Laboratory for Translational Medicine of Dermatology, Shenzhen Peking University, The Hongkong University of Science and Technology Medical Center, Shenzhen, Guangdong, China
#These authors contributed equally to this work and should be considered joint corresponding authors.
Prurigo nodularis is a highly pruritic and hyperplastic chronic dermatosis with unknown pathogenesis. Many pruritogenic mediators, including nerve growth factor, interleukin (IL)-31, thymic stromal lymphopoietin, and endothelin-1, are implicated in chronic itch and inflammation. This study investigated the mRNA levels and immunoreactivity of the nerve growth factor, IL-31, thymic stromal lymphopoietin, and endothelin axes in both lesional and perilesional skin in prurigo nodularis by using quantitative real-time PCR and immunohistochemistry studies. The nerve growth factor high-affinity receptor tyrosine kinase receptor A was upregulated while the low affinity receptor p75 neurotrophin receptor was downregulated in prurigo nodularis lesions. Downregulated expression of IL-31/IL-31 receptor A and endothelin-3/endothelin receptor B and upregulation of thymic stromal lymphopoietin receptor were found in prurigo nodularis lesions. Aberrant expression of nerve growth factor, IL-31, endothelin, and thymic stromal lymphopoietin axes was found in prurigo nodularis lesions, especially in the epidermis, indicating the importance of keratinocytes in prurigo nodularis pathogenesis.
Key words: prurigo nodularis; keratinocyte; itch; pruritogenic mediator; nerve growth factor.
Accepted Feb 26, 2019; E-published Feb 27, 2019
Acta Derm Venereol
Corr: Xia Dou, MD and Bo Yu, MD, Department of Dermatology, Peking University Shenzhen Hospital, Shenzhen, Guangdong 518036, China. E-mails: drdouxia@163.com; yubomd@163.com, and Yong Yang, MD, Department of Dermatology, Peking University First Hospital, Beijing 100034, China. E-mail: dryongyang@bjmu.edu.cn.
· Prurigo nodularis is a highly pruritic and hyperplastic chronic dermatosis with studies mainly focusing on cutaneous nerve fibres and neuropeptides.
· In this study, aberrant expression of nerve growth factor, interleukin-31, thymic stromal lymphopoietin, and endothelin axes was found in prurigo nodularis lesions, especially in the epidermis, indicating the importance of keratinocytes in prurigo nodularis pathogenesis.
· The nerve growth factor high-affinity receptor tyrosine kinase receptor A was upregulated while the low affinity receptor p75 neurotrophin receptor was downregulated in prurigo nodularis lesions, suggesting the importance of receptor imbalance in prurigo nodularis pathogenesis.
· These pruritogenic mediators may act as biomarkers for anti-pruritic and anti-inflammatory therapies of prurigo nodularis and need further studies.
Prurigo nodularis (PN) is a chronic skin disorder characterized by symmetrically distributed, highly pruritic and hyperkeratotic nodules (1, 2). Recently, PN was defined as a subtype of chronic prurigo that occurs due to neuronal sensitization to itch and the development of an itch-scratch cycle (3). It has been determined that PN has a heterogeneous origin, arising from dermatological (e.g., atopic dermatitis), systemic (e.g., chronic kidney failure) and neurological (e.g., brachioradial pruritus) or psychiatric diseases. However, it usually results from a combination of these different conditions or even from an unknown origin. Several reports have shown that many patients with PN show a predisposition to atopy (2, 4, 5). However, the pathogenesis of PN remains unclear, leading to limited effective treatment options (1).
The histopathological characteristics of PN are epidermal hyperplasia, inflammation and neurohyperplasia in the dermis, which suggests that complex interactions between keratinocytes, immune cells, and sensory neurons might occur and affect the formation and chronicity of PN. Keratinocytes might release chemokines/cytokines that modulate immune cell activity and vice versa. Both keratinocytes and immune cells can also produce pruritogens, algogens, and antinociceptive factors that act on sensory neurons, whereas sensory neurons release neuropeptides that target both keratinocytes and immune cells (6, 7).
To date, various pruritogenic molecules are implicated as mediators of chronic itch and inflammation, such as nerve growth factor (NGF), interleukin-31 (IL-31), thymic stromal lymphopoietin (TSLP), and endothelin-1 (ET-1) (6–8). Interestingly, these mediators can be produced by keratinocytes, thus suggesting that keratinocytes may play an important role in the pathogenesis of PN. However, studies of PN mainly focus on cutaneous nerve fibres and neuropeptides, with relatively scant attention to keratinocytes (1, 9).
In the current study, to assess the expression pattern of pruritogenic mediators in PN and to better understand the role of these mediators in PN pathogenesis, we investigated the expression of NGF, IL-31, TSLP, and ET axes in PN lesional and perilesional skin using quantitative real-time PCR and immunohistochemistry studies.
Patients with PN of Han Chinese ethnicity aged 18 years or older were enrolled. All patients were diagnosed with PN by two independent dermatologists according to clinical features and histopathological results. Clinically, PN is defined by the presence of numerous, itchy and symmetrically distributed, red-brown nodules. Use of systemic immunosuppressants or corticosteroids within the past 4 weeks, topical steroids or immunomodulators within the past 2 weeks, or moisturizers within the past 24 h before biopsy was restricted. Patients with skin infection or systemic disease were excluded. Patient clinical data were collected, including demographic data, serum total IgE level, and disease score via PRUNOSI (10), the verbal rating scale (VRS, scores range 0–3, 0 = none, 1 = mild, 2 = moderate, 3 = severe), visual analogue scale (VAS, scores from 0–10, 0 = no pruritus, 10 = worst imaginable pruritus) and dermatology life quality index (DLQI). Lesional skin and perilesional specimens from the extensor surface of patient extremities were obtained. The study was approved by the Ethical Committee of Peking University Shenzhen Hospital (No. 20160427) and was conducted according to the Declaration of Helsinki principles. Written informed consent was obtained from all participants.
Each skin specimen was equally divided into two parts (Fig. S1). One part, including the epidermis and dermis layers, was used to measure RNA expression via quantitative real-time PCR (qRT-PCR). The other part was used for haematoxylin and eosin (H.E.) and immunohistochemistry staining. H.E. staining was performed for all sections using standard procedures to confirm the diagnosis.
For immunohistochemistry, the sections were hydrated by passage through xylene and graded ethanol. After antigen retrieval for 17 min at 100°C in citrate buffer (pH 6.0), the slides were incubated in 3% H2O2 for 15 min to block endogenous peroxidase activity. Slides were blocked with 4% goat serum and 2% BSA for 1 h and then incubated with primary antibody (listed in Table SI) at 4°C overnight, followed by incubation with horseradish peroxidase (HRP)-labelled goat anti-rabbit IgG (1:1; Zhongshan Goldenbridge Co. Ltd., Beijing, China) for 45 min. All sections were counterstained using a DAB kit (MXB@, Biotechnologies, Fuzhou, China).
For all specimens, the staining intensity and extent were blindly evaluated by two independent dermatologists. Staining intensity was graded as follows: 1) none; 2) weak; 3) moderate; 4) strong. Staining extent was graded as follows: 1) 0–25%; 2) 25–50%; 3) 50–75%; 4) 75–100%. For each specimen, the staining index was assessed by multiplying the staining intensity score by the staining extent score, with the index range from 1 to 16 (11).
Total RNA was extracted from lesional and perilesional skin samples that included the epidermis and dermis layers using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) in accordance with the manufacturer’s protocol. RNA quality was confirmed with gel electrophoresis and a NanoDrop 2000 spectrophotometer (NanoDrop Technologies, USA), and then, the RNA was stored at –80°C until use. Reverse transcription of 2 μg of total RNA was performed using a GoScript™ Reverse Transcription System (Promega, Madisson, USA). qRT-PCR (40 cycles of denaturation at 95°C for 10 s, annealing at 60°C for 30 s, and extension at 72°C for 30 s) was performed using SYBR Green supermix (Bio-Rad, Hercules, CA, USA) on a CFX96 Real-Time PCR Detection System (Bio-Rad). The reference gene human β-actin (ACTB) was used to normalize the mRNA data. All reactions were performed in triplicate, and all tests were performed in triplicate. Fold differences in expression were calculated using the comparative cycle threshold (Ct) method by standardizing against ACTB expression. Quantification of target gene expression was performed using the 2→Ct method, with ΔCt being the mean Ct of ACTB subtracted from the mean Ct of the target gene. Gene expression levels were measured according to the above protocol, and the sequence-specific primers are listed in Table SII.
Statistical analysis was performed using SPSS (version 19.0) statistical software (SPSS Inc., Chicago, IL, USA). The clinical data are expressed as the means ± standard deviation (SD), while expression data are expressed as the means ± standard error of the mean (SEM). ATCB-normalized qRT-PCR expression values were log2-transformed before analysis. Paired Student’s t-test was used to compare differences in expression between lesional and perilesional skin. Mann-Whitney U test was used to compare differences in expression between atopic and non-atopic lesions. Spearman correlation was used to analyse associations among clinical markers and/or gene expression. A value of p < 0.05 was considered statistically significant.
A total of 11 patients with clinicopathologically confirmed PN (5 women and 6 men; mean age, 47.91 ± 12.48 years) were enrolled (Table I). Lesions of PN typically develop on the extensor surface of extremities, but an additional generalized distribution affecting the face, back, buttock or palms, as well as a “butter?y sign”, can be found (Fig. 1a–d). All lesional skin biopsies from subjects showed typical signs of PN. Specifically, pronounced hyperkeratosis, irregular acanthosis and pseudoepitheliomatous hyperplasia were observed in the epidermis. In the dermis, moderate lymphocytic inflammatory infiltrate and vertically oriented collagen bundles were observed (Fig. 1e).
Table I. General information of patients with prurigo nodularis
Fig. 1. Clinicopathological features of prurigo nodularis. (a–d) Nodules present on the extensor surfaces of the trunk, extremities and palms (with a “butterfly sign”). (e) Lesional skin biopsy showed pronounced hyperkeratosis, irregular acanthosis and pseudoepitheliomatous hyperplasia in the epidermis. In the dermis, there was moderate lymphocytic inflammatory infiltrate and vertically oriented collagen bundles (original magnification×100).
Approximately half of the patients had a history of atopy. Patients with past or present history of allergic diseases were defined as atopic PN, including AD, allergic conjunctivitis, allergic rhinitis and allergic asthma, whereas the rest of patients were non-atopic PN (5). Most of the patients presented with moderate to severe pruritus with an VRS score of 2 or 3, which is in accordance with a VAS score higher than 5. More than one pruritus sensation was mentioned by most patients, such as a combination of pruritus and stinging, burning and tingling. PRUNOSI scores ranged from 4 to 9 (mean 6.2 ± 1.7). The serum eosinophil counts were normal in all patients, and 7/11 patients showed an elevated total IgE level (> 200 IU/ml). A summary of patient characteristics is presented in Table I.
The biological effects of NGF are mediated through binding to the high affinity receptor tyrosine kinase receptor A (TrkA) and the low affinity receptor p75 neurotrophin receptor (p75NTR) (12). We first investigated the expression pattern of NGF-NGF receptors in PN lesional and perilesional skin. The mRNA levels of NGF and TrkA were significantly upregulated in lesional compared with perilesional skin (p < 0.05). However, in comparison with perilesional skin, the expression of p75NTR was significantly downregulated in PN lesional skin (p < 0.05) (Fig. 2a–c).
Fig. 2. Quantitative real-time PCR comparisons of selected pruritogenic mediators in lesional and perilesional skin are shown. (a–c) nerve growth factor (NGF) axis, (d–f) interleukin (IL)-31 axis, (g–i) thymic stromal lymphopoietin (TSLP) axis, and (j–n) endothelin (ET) axis (only 7 patients were tested for ET-1 and ET-2). Values show log2 expression/β-actin (ACTB) and are presented as the means ± SEMs. Stars above error bars denote comparisons with matched lesional and perilesional skin. *p < 0.05, **p < 0.01, and ***p < 0.001 vs perilesional skin determined by paired Student’s t-test. NL: perilesional skin; LS: lesional skin.
In immunostaining studies, both lesional and perilesional skin showed similar NGF immunoreactivity in the basal and spinous layers in the epidermis (Fig. 3a, b; Fig. 4a). Increased TrkA immunoreactivity was observed in the basal and spinous layers in lesional compared with perilesional epidermis (Fig. 3c, d; Fig. 4b). In PN lesional and perilesional skin, p75NTR staining was observed in all layers of the epidermis, with overall decreased staining in PN lesions (Fig. 3e, f; Fig. 4c).
Fig. 3. Representative images of immunohistochemistry staining of nerve growth factor (NGF) and interleukin (IL)-31 axes in lesional and perilesional skin are shown. (a, b) NGF, (c, d) tyrosine kinase receptor A (TrkA), (e, f) p75 neurotrophin receptor (p75NTR), (g, h) IL-31, (i, j) IL-31RA, (k, l) oncostatin M receptor (OSMR). (original magnification ×200; (e), original magnification ×100). NL: perilesional skin; LS: lesional skin.
Fig. 4. Staining quantification of selected pruritogenic mediators. Results are expressed as means ± SEMs. Stars above error bars denote comparisons with matched lesional (LS) and perilesional (NL) skin. *p < 0.05, **p < 0.01, and ***p < 0.001 vs perilesional skin determined by paired Student’s t-test.
IL-31 is a potent pruritogenic cytokine that signals through a heterodimeric receptor complex comprising IL-31 receptor A (IL-31RA) and oncostatin M receptor (OSMR) (13). We next investigated the expression pattern of IL-31 and the IL-31 receptor. At the mRNA level, we found a significant decrease in IL-31 and IL-31RA expression in lesional compared with perilesional skin (p < 0.05). No significant difference was found in OSMR expression (Fig. 2d–f).
Based on immunostaining, sparse IL-31 staining was found in the basal and spinous layers in lesional and perilesional epidermis (Fig. 3g, h; Fig. 4d). IL-31RA staining was mainly observed in the suprabasal epidermis and was decreased in lesional compared with perilesional epidermis (Fig. 3i, j; Fig. 4e). OSMR showed a moderate homogenous immunoreactivity in the epidermis in all samples analysed (Fig. 3k, l; Fig. 4f).
TSLP signals through TSLP receptor, which is composed of TSLP receptor (TSLPR) and IL-7 receptor α chain (IL-7RA) (14). For the TSLP axis, we found a significant increase in only the TSLPR mRNA levels in lesional compared with perilesional skin (p < 0.05). No significant differences were found in TSLP and IL-7RA expression (Fig. 2g–i).
In immunostaining studies, moderate TSLP staining was found in all layers of lesional and perilesional epidermis (Fig. 4g; Fig. 5a, b ). TSLPR was mainly observed in the basal and lower spinous layers in all samples, with more TSLPR-positive cells in lesional than in perilesional epidermis (Fig. 4h; Fig. 5c, d). Strong IL-7RA staining was found in all layers of lesional and perilesional epidermis (Fig. 4i; Fig. 5e, f).
Fig. 5. Representative images of immunohistochemistry staining of thymic stromal lymphopoietin (TSLP) and endothelin (ET) axes in lesional and perilesional skin are shown. (a, b) TSLP, (c, d) TSLPR, (e, f) interleukin (IL)-7RA, (g, h) ET-3, (i, j) ETA, (k, l) ETB. (original magnification ×200). NL: perilesional skin; LS: lesional skin.
The ET axis is composed of the 3 endogenous ligands ET-1, ET-2, and ET-3 and ET receptors A and B (ETA, ETB) (15). For ET axis expression, we found significantly downregulated mRNA levels of ET-1, ET-2, ET-3, and ETB in lesional compared with perilesional skin (p < 0.05), especially for ET-3 and ETB (Fig. 2j–n).
In immunochemistry studies, ET-3 staining was mainly observed in the suprabasal layers of lesional and perilesional skin, with decreased staining in lesional epidermis (Fig. 4j; Fig. 5g, h). Both ETA and ETB showed cytoplasmic staining patterns. Similar immunoreactivity was found for ETA in all layers of the epidermis in lesional and perilesional skin samples (Fig. 4k; Fig. 5i, j). Decreased staining was observed for ETB in lesional compared with perilesional epidermis (Fig. 4l; Fig. 5k, l).
We investigated the lesional expression of these mediators in 5 atopic patients and 6 non-atopic patients, and no differences were found for any pruritogenic mediators (Fig. S2). Then, we compared lesional and perilesional expression in the 5 atopic patients. Higher expression of TrkA (p = 0.087) and lower expression of p75NTR (p = 0.079) were observed in lesions (Fig. S3b, c). Significantly lower IL-31RA, ET-3 and ETB levels were found in lesional vs perilesional skin (Fig. S3e, j, l). For the 6 non-atopic PN patients, increased NGF and decreased p75NTR mRNA were found in lesional skin. We also found significant increase of IL-31, decrease of TSLPR and ET-3/ETB mRNA levels in lesional vs perilesional skin (Fig. S4).
We finally evaluated associations among different clinical measures (age, duration, PRUNOSI, VRS, VAS, DLQI, and total IgE) and correlations between these clinical markers and gene expressions in PN lesions. DLQI was positively associated with PRUNOSI, VRS and VAS (p < 0.05) (Fig. S5). Moreover, positive correlations were found between lesional NGF mRNA levels and age (p < 0.05) and a trend between lesional NGF mRNA levels and PN disease duration (Fig. S5).
It has been shown that neuropeptides/neurotrophins and cutaneous nerve fibres play an important role in the pathogenesis of PN (9). NGF is a neurotrophic factor synthesized by various cell types, including keratinocytes. In addition to activation, sensitization and sprouting of skin nerves, NGF is also important in proliferation and differentiation of keratinocytes acting via NGF receptor (9, 11, 16). TrkA is considered an NGF-specific receptor due to its greater affinity and specificity for NGF, while p75NTR can also bind other neurotrophins (16).
Previous studies of PN have focused on the lesional dermis and showed strong NGF, TrkA and p75NTR staining in hyperplastic nerve fibres in lesional dermis, which may contribute to dermal neurohyperplasia, neurogenic inflammation and itch induction (9, 11, 17, 18). In this study, we found increased TrkA immunostaining in lesional epidermis. Another study found that some PN patients show increased TrkA staining in lesional epidermis, and in vitro studies showed that Th2 cytokines enhanced TrkA expression and upregulated the proliferation of keratinocytes (11). Therefore, TrkA might play an important role in epidermal hyperplasia in PN.
Here, a remarkable downregulation of p75NTR was found in PN lesions, which is also the case in AD and psoriatic lesional epidermis (11, 19, 20). Acting as a stranger in the TNFR superfamily (12), p75NTR is linked to apoptosis and carcinogenic processes in the skin (19, 21–23). A lack of p75NTR in psoriatic transit-amplifying cells leads to reduced keratinocyte apoptosis, which plays an important role in the maintenance of epidermal homeostasis (19). In the nervous system, opposing signalling effects of tropomyosin receptor kinases (Trks, including TrkA) and p75NTR have been reported, with Trk receptors mediating neurotrophin-enhanced cell survival and p75NTR promoting cell death of sensory and sympathetic neurons (16). Similarly, in PN, we speculate that NGF-TrkA upregulation might mainly contribute to keratinocyte proliferation, while p75NTR downregulation might reduce apoptosis of keratinocytes.
IL-31 is a potent pruritogenic cytokine that is preferentially expressed by CD4+ T helper 2 (TH2) cells and skin-homing CD45RO+CLA+T cells. In PN, it has been reported that IL-31 mRNA is increased in PN lesions compared with normal skin or may be higher in lesional than in non-lesional skin in PN patients (24–26). In this study, we found downregulation of IL-31 mRNA in PN lesions. Although no significant differences were found, we observed higher expression of IL-31 mRNA in lesions from the 5 atopic patients than in those from the 6 non-atopic patients (Fig. S21). These results suggest a much more important role of IL-31 in atopic vs. non-atopic PN patients, and thus, further studies are needed (24).
IL-31 signals through IL-31 receptors composed of IL-31RA and OSMR, which can be expressed in keratinocytes (13, 27). In a previous study, epidermal staining for IL-31RA and OSMR yielded moderate homogenous immunoreactivity in PN keratinocytes (28). In this study, we found decreased IL-31RA mRNA and staining in PN lesions. It is worth noting that IL-31/IL-31RA signalling has been shown to inhibit proliferation and deregulate differentiation in both primary normal human epidermal keratinocytes (NHEKs) and HaCaT cells (29, 30). Decreased IL-31RA expression during the process of differentiation leads to a loss of IL-31 responsiveness, which might lead to uncontrolled proliferation and decreased apoptosis of keratinocytes (29). Therefore, downregulation of IL-31/IL-31RA might be involved in epidermal hyperplasia in PN.
TSLP is one of the most important mediators in the pathogenesis of AD. It is mainly produced by epithelial cells and signals through IL-7RA and the higher affinity receptor TSLPR. The pathophysiologic profile of TSLP includes itch induction and allergic inflammation (14, 31). Overexpression of TSLP has been reported in keratinocytes of AD patients and in tape-stripped barrier-disrupted skin (31, 32). In this study, although scratching-induced epidermal barrier defects were frequently found in PN patients, aberrant TSLP expression was not found between lesional and perilesional skin.
TSLP released from keratinocytes can act directly on a subset of TSLPR(+)/TRPA1(+) sensory neurons to trigger robust itch in AD patients (31, 32). Moreover, increased epidermal TSLP and TSLPR have been reported in scars with post-burn pruritus compared with scars without pruritus or normal tissues. Additionally, an enhanced number of epidermal PGP 9.5-positive nerve fibres has also been observed in scars with post-burn pruritus, suggesting that higher expression of TSLP-TSLPR in epidermis is related to neurohyperplasia in post-burn scars (33). In PN lesions, no intraepidermal neurohyperplasia has been reported, but intraepidermal neurohypoplasia has been observed (18, 26, 34). Here, we found increased TSLPR expression in lesional basal and spinous layers. Whether TSLP-TSLPR signalling plays a role in PN pathogenesis, including in the proliferation of keratinocytes or itch induction, needs further study.
Endothelins (ETs) are a family of small, vasoactive, 21-amino acid peptides referred to as ET-1, ET-2 and ET-3 that act by activating two G protein-coupled receptors, ETA and ETB (15). Regarding the affinity of the 3 isoforms to ET receptors, ET-1 and ET-2 are more potent than ET-3 (ET-1 = ET-2>ET-3) for ETA, and all 3 isoforms are equally effective for ETB (15). ETs and their receptors are expressed in a variety of human tissues and cell types (e.g., keratinocytes). ET-1 primarily plays an important role in itch induction (35, 36) and inflammatory responses (35–39). In this study, we found decreased ET-1 mRNA levels in PN lesional skin. One study has shown enhanced ET-1 staining in PN lesional epidermis compared with normal skin, which induces pruritus through ET-1/ETA/ECE-1/ERK1/2 signalling (36). Here, we also found downregulation of ET-2 and ET-3 in PN lesional skin. However, the role of ET-2 and ET-3 in inflammation and itching pathogenesis remains unknown. For ET receptors in this study, no difference in ETA but a remarkable decrease in ETB expression was found in PN lesional skin. A previous study showed that activation of ETB expressed in keratinocytes exerts antipruritic effects on ET-1-induced itch via peripheral κ-opioid receptors (35). Thus, decreased ETB in PN lesions might lead to impaired antipruritic function in PN lesions.
From correlation analysis, we found that only NGF was positively correlated with age and disease duration at the lesional skin level, suggesting a role of NGF in the ongoing formation and evolution of PN. Moreover, severe itching has a large negative impact on quality of life in PN patients, suggesting an urgent need to break the itching-scratching cycle with effective PN treatments.
In summary, we preliminarily identified the expression patterns of pruritogenic mediators, including the NGF, IL-31, TSLP and ET axes, in PN lesions. Aberrant expression of these factors was found in lesional skin, especially in keratinocytes, indicating the importance of keratinocytes in the pathogenesis of PN. For epidermal hyperplasia in PN lesions, increase of NGF-TrkA, decrease of p75NTR and IL-31/IL-31RA may make a contribution through upregulating proliferation and reducing apoptosis of keratinocytes, while decreased ETB in PN lesions might lead to impaired antipruritic function in PN lesions. However, the exact role of TSLP-TSLPR signalling in PN pathogenesis remains unclear. All these results are in the basis of small sample size and need further studies.
We thank all the patients and their family members for participating in this study. This work was supported by grants from the National Natural Science Foundation of China (81472878, 81502727, 81673053) and Shenzhen Research Grant (JCYJ20160428173958860, JCYJ20170411090739316, JCYJ20170306161807726).
The authors have no con?icts of interest to declare.


 Click to show fullsize
Click to show fullsize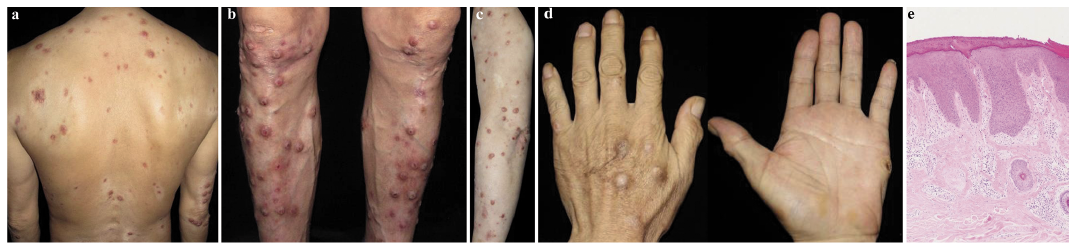 Click to show fullsize
Click to show fullsize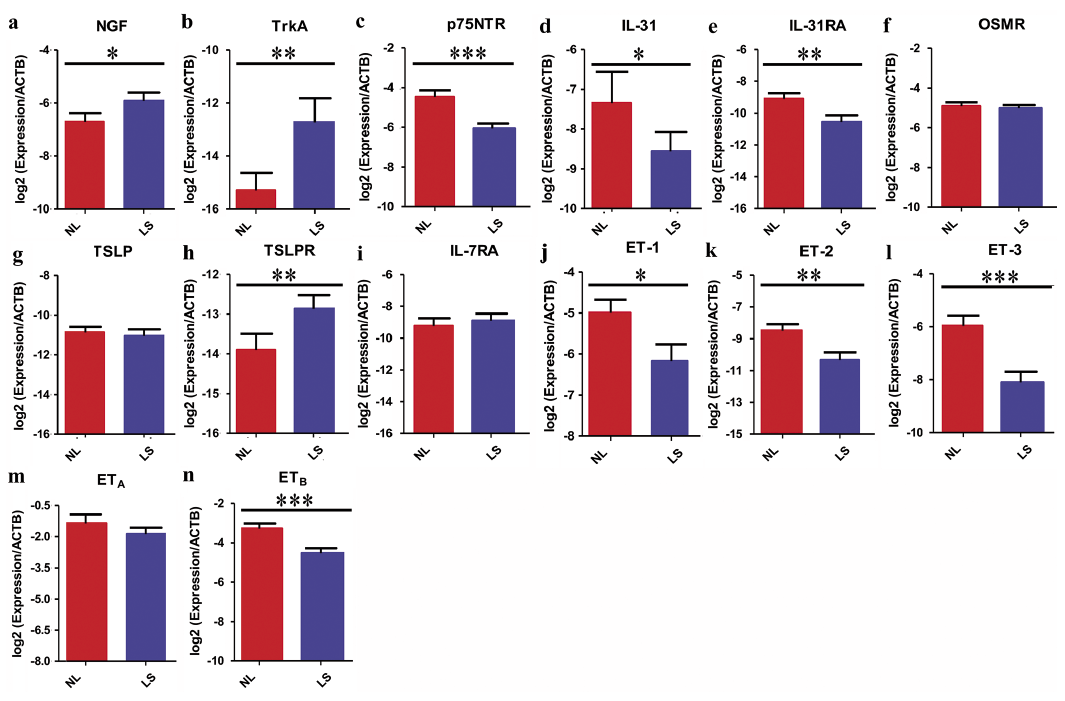 Click to show fullsize
Click to show fullsize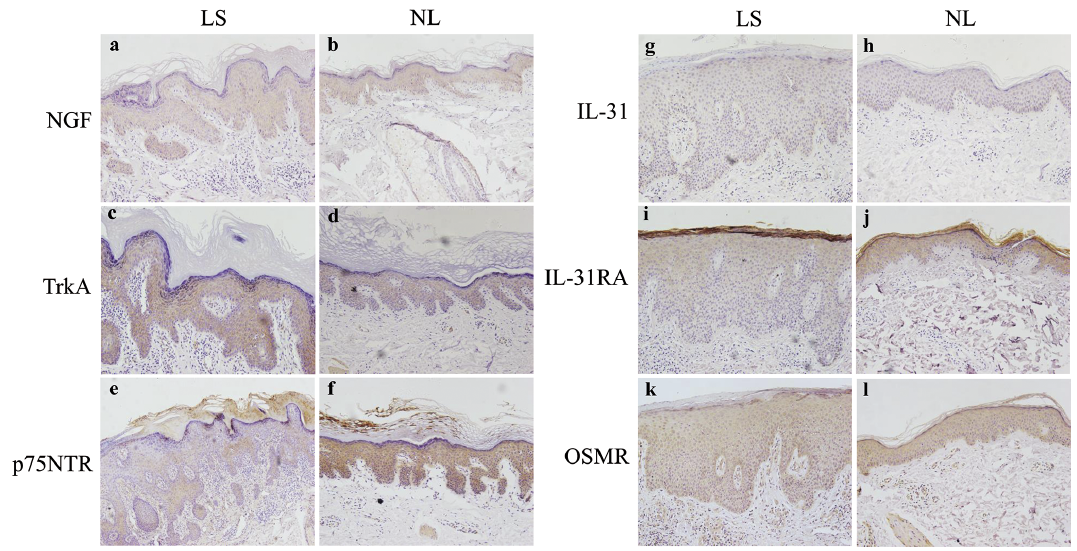 Click to show fullsize
Click to show fullsize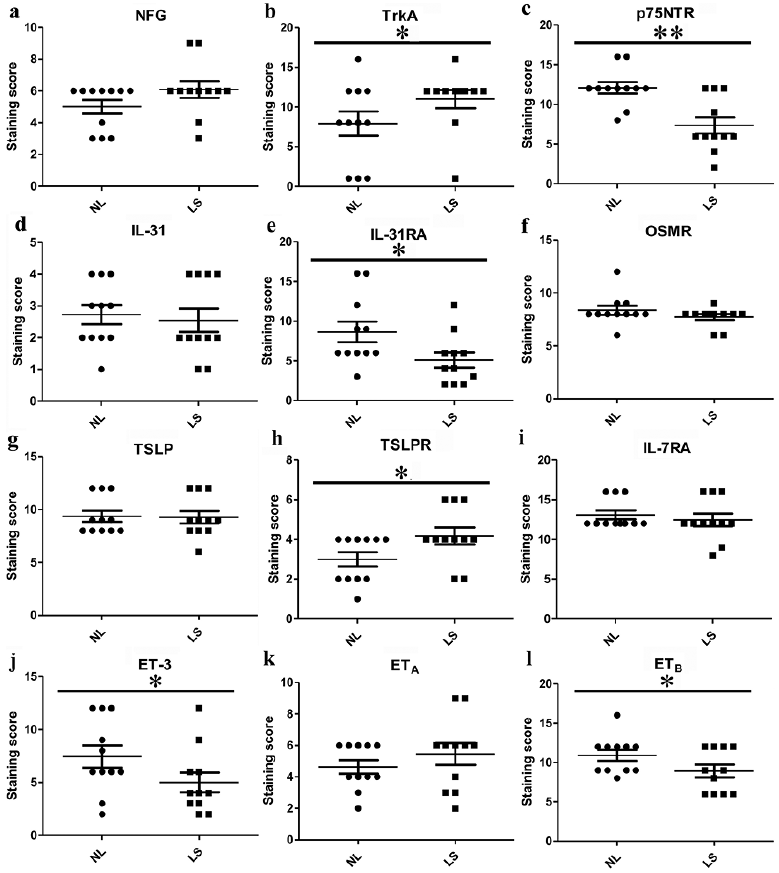 Click to show fullsize
Click to show fullsize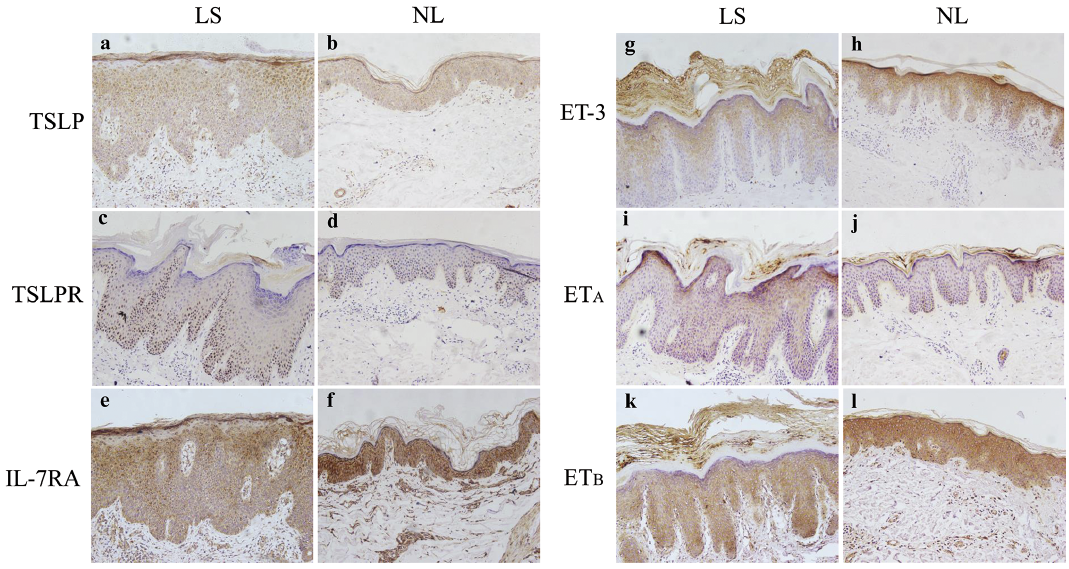 Click to show fullsize
Click to show fullsize