


1Juntendo Itch Research Center (JIRC), Institute for Environmental and Gender Specific Medicine, 2Anti-aging Skin Research Laboratory, Juntendo University Graduate School of Medicine, and 3Department of Dermatology, Juntendo University Urayasu Hospital, Chiba, Japan
Chronic itch is a burdensome clinical problem that often accompanies pathological dry skin-based conditions, such as atopic dermatitis, and systemic disorders, such as kidney diseases, with an unclear pathomechanism and treatments. One of the basic mouse models to investigate mechanisms of itch associated with dry skin is a mixture of acetone and ether followed by water. Animal studies using the acetone and ether followed by water model have revealed that many mediators and receptors, e.g. mas-related G protein-coupled receptor family, transient receptor potential, and chemokines, are responsible for itch and its hypersensitivity, supporting the hypothesis that dry skin-induced itch is a histamine-independent pathway. New insights have been acquired into the interplay between neurones and non-neuronal cells in the initiation, modulation, and sensitization of itch. Several therapeutic options for itching have thus been developed. This review summarizes the updated pathogenesis and therapeutic strategies for itch in dry skin conditions.
Key words: dry skin; hypersensitivity; itch; sensory neuron; mouse model.
Accepted Oct 15, 2019; Published Jan 9, 2020
Acta Derm Venereol 2020; 100: adv00024.
Corr: Mitsutoshi Tominaga, Juntendo Itch Research Center (JIRC), Institute for Environmental and Gender Specific Medicine, Juntendo University Graduate School of Medicine, 2-1-1 Tomioka, Urayasu, Chiba 279-0021, Japan. E-mail: tominaga@juntendo.ac.jp
Itch is an unpleasant sensation that may disturb quality of life, and for which the pathomechanism and appropriate treatments are unclear. Chronic itch, which lasts more than 6 weeks, often accompanies pathological dry skin-based conditions, such as xerosis, atopic dermatitis, liver and kidney diseases. A decline in skin barrier function is thought to be the primary cause of itch induced by dry skin. Many kinds of mediators, receptors, and channels are involved in itch signalling among the skin nervous system, skin cells, and central nervous system. Several therapeutic options for itching have thus been developed, such as phototherapy, phospholipids, antioxidants, and emollients.
Skin, the body’s largest organ, serves as a first physiological barrier against the external environment. The barrier function of the skin is exerted by the epidermis, the most superficial layer of the skin, of which the stratum corneum (SC) is largely responsible for the barrier function. There are 2 elements important for the maintenance of SC humidity: intercellular lipids, which form the main barrier against diffusion of water across the SC, and natural moisturizing factor, which has a key role in the absorption of water in the SC. Impaired skin barrier integrity causes excessive water loss and leads to skin dryness (1, 2).
Dry skin is characterized by a scaly, rough, cracked, and fissured surface, and is closely associated with the somatosensory sensation of itch, especially chronic itch (3). Dry skin with chronic itch is the most common clinical manifestation of dermatoses, such as xerosis, atopic dermatitis (AD), and psoriasis, and is a common cutaneous manifestation in pruritic systemic diseases, such as chronic kidney disease (CKD), chronic liver diseases (CLD), and diabetes mellitus (DM) (4).
Histamine is a well-known substance that induces itch; however, antihistamines (histamine H1-receptor blockers) are not fully effective in many dermatological and systemic diseases characterized by dry skin, suggesting that dry skin is an important feature of antihistamine-resistant (histamine-independent) itch (2). The underlying condition of dry skin is impaired function of the skin barrier, which can be caused by environmental factors, such as sun exposure, temperature, humidity, and genetic factors, such as filaggrin mutations (1, 5, 6). To assess skin barrier func-tion, transepidermal water loss (TEWL), SC hydration, and pH are commonly used (1). The signs and clinical manifestations of dry skin are not only physically uncomfortable, but also affect patients psychologically (7).
Disease-related dry skin
Aged skin. Xerosis is one of the most prevalent dry skin conditions in the aged population worldwide (8), affecting over 50% of individuals aged ≥ 65 years (9). Multiple skin changes in the elderly are related to xerosis: (i) alterations in the barrier function of SC, including cellular and intercellular lipid matrix changes; (ii) pH variations; (iii) alterations in SC proteases; (iv) reduced activity of sebaceous and sweat glands; and (v) decreased oestrogen levels. All of these factors may lead to itch induction (10).
Inflammatory skin diseases. Dry skin itch is a common symptom in dermatoses characterized by dysfunction of the skin barrier, such as AD and psoriasis. In these diseases, pruritogens, such as cytokines and chemical mediators, are released from the affected area (5, 11). Pruritogens induce itch mainly by acting on the sensory nerves, and the affected area is scratched, then further aggravates dermatitis (12). This vicious cycle is called the “itch-scratch cycle”. Skin hyperesthesia (a skin condition that involves an abnormal increase in sensitivity to stimuli) occurs in inflammation, such as AD (5). Elongation of the sensory nerve in the epidermis to immediately underneath the SC, due to drying and inflammation, is considered to be a cause of skin hyperesthesia. Nerve growth factor (NGF), amphiregulin (AR), and artemin (ARTN), which are nerve elongation factors (NEFs), and semaphorin 3A (Sema3A, a nerve repulsion factor (NRF)), are related to this aberrant nerve elongation and sprouting in AD (13).
More recently, Pogatzki-Zahn et al. (14) reported skin hyperesthesia in patients with chronic pruritus, such as AD, but it was not related to hyperinnervation in the epidermis, observed as a decreased number of cutaneous nerves crossing the basement membrane. The authors speculated that, although the nerves crossing the basement membrane were reduced, increased intraepidermal sprouting of nerves is possible. Another possibility is that the density, structure, and functional properties of intraepidermal nerves fluctuate in different skin disease states, especially in acute and chronic phases.
Systemic diseases. Dry skin is also a common cutaneous manifestation in pruritic internal diseases, such as CKD, CLD, and DM (4). Skin dryness may appear at different stages of CKD, but it is more frequently diagnosed in dialysis subjects (45%) (15). The functional abnormalities of eccrine sweat glands may account, at least in part, for dry skin in uraemic patients (16). It has been suggested that dry skin can cause itch in CKD; however, objective measurements of the barrier function of the skin, such as the degree of hydration of the skin, lipid bilayer abnormalities, and dryness of the skin, do not always correlate with pruritus (17).
The pathogenesis of pruritus in CLD is poorly understood and often refractory to treatment, with a prevalence of 40.3% (18). Several potential itch-causing substances may be involved, including bile salts, endogenous opioids, histamine, serotonin, and steroids (19). We reported recently that the plasma dynorphin A level of endogenous opioids correlates with the severity of pruritus and may reflect its degree in patients with CLD (20).
Skin disorders are common complications and comprise a broad spectrum of disorders in both type 1 and type 2 DM, e.g. cutaneous infection, dry skin, and pruritus (21). Clinical observations are supported by a reduced hydration of the SC and reduced sebaceous gland activity in patients with DM. Even in the absence of clinically apparent dry skin, patients with diabetes have an impaired desquamation process (22). Pruritus is more common in patients with diabetes who have dry skin or diabetic neuropathy (21). Higher postprandial glucose levels were reported to result in a higher probability of having generalized pruritus (23).
Dry skin mouse models
Acetone-treated model (acute dry skin model with no itch). One mouse model to induce dry skin uses acetone application. The hair of mice is shaved over the rostral part of the back at least 3 days before acetone treatment. The shaved area was treated with acetone-soaked cotton balls for 5 min. In the control group, the shaved area was treated with sterile water (3, 24).
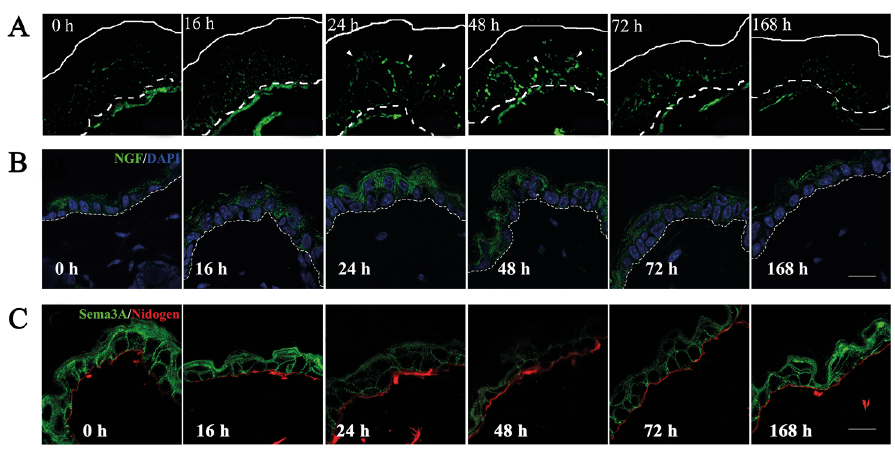
Analyses of experimental animals treated with acetone demonstrated that intraepidermal innervation-related factors, such as NGF and ARTN gene expression, were increased in the epidermis, and the artificial restoration of the barrier immediately following barrier disruption by acetone treatment inhibited the increase in these mRNA levels (3, 24). Others observed the release of histamine from mast cells in the skin of acetone-treated mice (25). We found that acetone-treated mice displayed a rapid increase in TEWL and a decrease in SC hydration during the first hour after treatment, which returned to normal by 48 h after the treatment. Thus, the acetone-treated mice manifest the characteristics of dry skin and have altered cutaneous barrier permeability. No scratching behaviours or epidermal hyperplasia were observed in the acetone-treated mice, although there was an increase in nerve fibre density in the epidermis (Fig. 1A). Of note, we found that the expression of epidermal NGF and AR (which promote nerve growth) was increased (3), but Sema3A (which inhibits nerve growth) expression was decreased (Tominaga et al., unpublished data) before the penetration of nerve fibres into the epidermis (Fig. 1B and C). The increase in intraepidermal nerve fibres may be an important factor for the regulation of itch in dry skin (3).
Acetone/ether/water (AEW)-treated model (chronic dry skin model with itch). The AEW-treated mouse model is one of the most well-known mouse models for the study of dry skin-induced itch (26). The hair of mice was shaved over the rostral part of the back at least 3 days before the start of the experiment. To disrupt the cutaneous barrier, cotton (2 × 2 cm) soaked in a mixture of acetone and ether (1:1) was placed on the shaved area for 15 s. Immediately after AE treatment, cotton soaked with distilled water was placed on the same area for 30 s. Treatments were performed twice daily under ether anaesthesia for 5–7 consecutive days. TEWL and scratching behaviour were increased, and SC hydration was decreased, under this treatment. The histopathological analysis showed that the AEW-treated mice had marked epidermal hyperplasia, parakeratosis, and infiltration of nerve fibres into the epidermis, but no infiltration of inflammatory cells in the dermis (26, 27). Overall, the AEW treatment produces marked skin barrier dysfunction, robust scratching, and changes in gene expression in sensory nerves and the skin (26), which recapitulate the dry skin symptoms present in many chronic itchy conditions in humans (28). There was no apparent difference in AEW-induced spontaneous scratching between mast cell-deficient mice (WBB6F1-W/WV) and normal litter-mates (WBB6F1-+/+) (26).
This dry skin model also exhibits alloknesis (scratching behaviour evoked by a stimulus that is normally non-pruriceptive) and hyperknesis (the abnormal pruriceptive state, in which a normally pruritic stimulus elicits a greater than normal duration and/or magnitude of itch) (27, 29), as described later in this review.
Special diet food model. HR-1 hairless mice fed a special diet (HR-AD) is one of the dry skin-based experimental mouse models. Mice were fed HR-AD for 48 days. These mice exhibited severe dry skin symptoms accompanied by a decrease in skin water-holding capacity, increase in TEWL, and prolonged scratching bout duration. Marked epidermal hyperplasia, and increase in circulating T cells and serum IgE are observed (30). Lipid composition analysis revealed that HR-AD is an essential fatty acid (EFA)-deficient diet. Feeding HR-AD with EFA inhibits the symptoms of dry skin (31). EFA deficiency was reported to depress skin barrier function due to structural changes in ceramides, and reduced elaboration and deposition of epidermal intercellular lipids (32). Therefore, HR-AD causes deterioration of the skin barrier function due to EFA deficiency.
Fig. 1. Alterations in nerve fibre distribution, and nerve growth factor (NGF) and Sema3A expression in the epidermis of acetone-treated dry skin model mice. (A) Sequential alteration of intraepidermal nerve growth in acetone-treated mice was examined by immunohistochemistry using an anti-PGP9.5 antibody. (B, C) Maximum expression of NGF (green) was noted 16–24 h after the treatment (B). In contrast, the expression level of Sema3A (green) was decreased 24 h after acetone treatment (C). These expression levels gradually returned to normal by 168 h after the treatment. Nuclei are counterstained by DAPI (blue). The broken lines in panel A indicate the border between the epidermis and dermis (basement membrane). The basement membrane in panel B was stained with an anti-nidogen antibody (red). White and broken lines indicate the skin surface and the border epidermis and dermis (basement membrane), respectively. Arrowheads indicate epidermal nerve fibres (green). Scale bars: 15 μm.
The sensation of itch is generated by the binding of itch-inducing substances to their cognate receptors on peripheral sensory afferents, e.g. unmyelinated C-fibre afferents and thinly myelinated Aδ-fibre afferents. The evoked action potential is transmitted through the ascending sensory pathway to the somatosensory cortex, resulting in the perception of itch (Fig. 2).
Fig. 2. Mechanisms and management of dry skin-induced itch. The perception of itch starts when endogenous and exogenous itch mediators activate their respective receptors/channels expressed in peripheral sensory afferents. Electric signals generated in the peripheral nerve endings are transmitted to the somatosensory cortex in the brain through the spinal cord, resulting in the recognition of itch. (A) In healthy skin, Sema3A, a nerve repulsion factor (NRF) produced mainly by keratinocytes (KCs), is dominant. It maintains the cutaneous nerve fibres under the dermo-epidermal junction. (B) During environmental stimuli in acute dry skin conditions, nerve growth factor (NGF), an epidermal nerve elongation factor (NEF) produced by KCs, is prominent and induces the elongation of cutaneous nerve fibres into the epidermis. This elongation may also be affected by thymic stromal lymphopoietin (TSLP) and interleukin (IL)-33 released from KCs. NGF also promotes matrix metalloproteinase (MMP)-2 and MMP-8 production in sensory nerve fibres, which leads to the penetration of nerve fibres into the basement membrane and their growth. Emollients and phospholipids are effective at alleviating the symptoms in this phase. (C) In chronic dry skin accompanying the itch-scratch cycle, such as in systemic or inflammatory skin diseases, more sensory nerve fibres penetrate the epidermis. In addition to substances released from KCs, the non-peptidergic C-fibres (NP cluster) are also involved in itch signalling, along with astrocytosis in the spinal cord. More treatments have been confirmed to be beneficial in this condition.
Sensory neurones
Nerve elongation and repulsion factors. In healthy skin, most cutaneous nerve fibres terminate under dermoepidermal junctions. An increased intraepidermal nerve density has been observed in the skin of patients with pruritic dermatological diseases, such as senile xerosis and AD (13), as well as in dry skin mice models (3, 33). The controlling mechanism of cutaneous nerve density is regulated by the balance of NEFs, such as NGF, ARTN and AR, and NRFs, such as Sema3A, produced by keratinocytes (5, 13). These axonal guidance molecules may also act on keratinocytes, immune cells, and vascular endothelial cells, and may be indirectly involved in the regulation of itching (34).
Matrix metalloproteinase (MMP)-2 and MMP-8. The process of cutaneous nerve growth in dry skin requires several MMPs for growth cones to penetrate the 3-dimensional extracellular matrix (ECM) barriers (Fig. 2). Using in vitro models of ECM, we found that MMP-2 localized on the growth cone functioned in penetration into the basement membrane (35). In addition, MMP-8 secreted by nerve fibres was reported to be involved in nerve growth within the dermis (36). The levels of expression of MMP-2 and MMP-8 were upregulated by NGF and down-regulated by Sema3A. The selection and up-regulation of MMPs corresponding to the ECM components surrounding the growing nerve fibres may be required for efficient nerve fibre penetration, suggesting that the coordinated activation of neurotrophin and ECM-integrin signalling is necessary for efficient and long-distance axon extension (37). As class 3 semaphorin signalling inhibits integrin-mediated adhesion signalling, Sema3A stimulation of growing nerve fibres may provide a reverse signalling pathway for these events (38).
Peptidergic fibres. Substance P (SP) and calcitonin gene-related peptide (CGRP) are neuropeptides produced by sensory nerves in the dermis to communicate with different cell populations in the different layers of the skin, which in turn stimulate nerve fibres. An increase in the elongated epidermal peripheral nerve fibres consist of SP/CGRP-containing C-fibres, which usually represent epidermal peptidergic nerve fibres, has been reported in the AEW model (39, 40).
The neuropeptide gastrin-releasing peptide (GRP) is characterized as a neurotransmitter that specifically relays itch signals and specifically expressed in a small subset of peptidergic dorsal root ganglion (DRG) neurones. Genetic ablation of GRP receptor (GRPR)+ neurones resulted in significant reduction in the scratching response to multiple pruritogens (41). The transcription factor Tlx3 in the spinal cord was demonstrated to be essential for the development of GRPR+ neurones (42). Huang et al. (43) reported that Tlx3 conditional knockout (Tlx3F/F; Nav1.8-Cre mice specifically lost Tlx3 expression in most TrkA-lineage DRG neurones) mice scratched much less compared with controls in the dry skin model, suggesting impairment of dry skin-induced chronic itch in these mice.
Non-peptidergic fibers. C-fibres have been divided into peptidergic and non-peptidergic subsets mainly on the basis of neurochemical criteria. The peptidergic neurones are mostly marked by neuropeptides, including SP and CGRP, whereas non-peptidergic neurones are commonly labelled by the purinergic P2X3 receptor and the plant lectin isolectin B4 (IB4) (44). On the contrary to previous reports of SP and CGRP involvement in the AEW model, a recent study reported that AEW treatment increased non-peptidergic intraepidermal fibres, but not CGRP+ fibres, suggesting that a specific subset of non-peptidergic fibres function in dry skin itch (45), as is observed in itch behaviour of the imiquimod-induced psoriasis mouse model (46).
Protease-activated receptors (PARs). PARs consist of 4 members: PAR-1, PAR-2, PAR-3, and PAR-4. PARs other than PAR-3 are expressed in cutaneous nerve fibres, keratinocytes, mast cells, and macrophages, and are considered involved in itch (27, 47). Spontaneous scratching behaviour in dry skin-treated animals was significantly attenuated by a PAR-2 antibody either delivered locally to the dry skin area or systemically. In addition, DRG cells from AEW-treated mice exhibited significantly larger responses to the PAR-2 agonist, implicating a role for endogenous agonists of this receptor in chronic itch (27).
Mas-related G protein-coupled receptor family (Mrgpr). The Mrgpr family in mice can be grouped into several subfamilies: MrgprA, MrgprB, MrgprC, and MrgprD-G (48). MrgprA3, MrgprC11, and MrgprD in mice, which are expressed only on small-diameter sensory neurones in the DRG and trigeminal ganglia (TG), and were recently suggested to be involved in the transmission of itch (49–51). The expression of mRNAs encoding MrgprA3 and MrgprC11 was found to be higher in AEW-treated dry skin model mice than in water-treated controls (52). Moreover, the ablation of MrgprA3+ DRG neurones reduced chronic itch induced by AEW treatment, suggesting that MrgprA3 functions in dry skin-related itch (53). The increases in expression of MrgprA3 and MrgprC11 were inhibited in acid-sensing ion channel 3 (ASIC3) knockout mice (52), suggesting that fluctuations in skin pH are involved in dry skin-related itch.
Transient receptor potential family (TRP). The TRP channels are known as polymodal cellular sensors. The ion channel TRP subfamily A member 1 (TRPA1) was previously reported to mediate acute histamine-independent itch, e.g. sensory neurone activation and itch behaviour downstream of 2 histamine-independent pruritogens, chloroquine and BAM8-22 (49, 54). Wilson et al. (55) found that functional TRPA1 is required for the dry-skin-evoked phenotypes, including AEW-evoked scratching, epidermal hyperplasia, and expressional changes in the skin. Among the human disease genes, TRPA1 regulates both scratch-dependent and scratch-independent changes: AQP3, IL-33, chemokine receptor CXCR2, lipocalin, Slc9a3r1, and S100A9 require TRPA1, and are independent of the itch-scratch cycle, whereas CCL27 and Tenascin C (TNC) are scratch- and TRPA1-dependent. These genes play diverse roles in the initiation and maintenance of chronic itch (55).
TRP cation channel subfamily V member 1 (TRPV1) is a heat-sensitive cation channel that is selectively expressed in a population of primary sensory neurones in TG and DRG, which plays an important role in thermal and pain sensations (56). Yu et al. (28) reported an increased innervation density of TRPV1-expressing sensory fibres in the skin of AEW model mice due to expansion of this channel. This may also be partly involved in the induction and/or enhancement of itch in dry skin.
NP clusters of sensory neurones. Usoskin et al. (57) reported 4 neuronal clusters (further divided into 11 fundamentally distinct types of sensory neurones) in single cells of sensory neurones from mouse lumbar DRGs. The first cluster is the NF cluster (including NF1-5), which expresses neurofilament heavy chain (Nefh) and parvalbumin (Pvalb), and was previously associated with myelinated DRG neurones. The second, the PEP cluster (including PEP1-2), expressed SP (Tac1), TRKA (Ntrk1) and CGRP, which were previously associated with peptidergic nociceptors. The third, the NP cluster (including NP1-3), expressed Mrgprd and P2rx3, which were previously associated with non-peptidergic nociceptors. The fourth, the TH cluster, exhibited distinct expression of tyrosine hydroxylase (Th) and has been described in a distinct subclass of unmyelinated neurones. Furthermore, NP1, NP2, and NP3 neuronal types were reported to function in itch, and NP3 is likely to sense and transduce inflammatory itch. They detected lysophosphatidic acid–responsive neurones (Lpar3 and Lpar5) in the NP1 class, chloroquine-responsive neurones (Mrgpra3 and Mrgprx1) in NP2, and interleukin (IL)-31 (Il31ra and Osmr)- and cysteine leukotriene (Cysltr2)-responsive neurones, neuropeptides natriuretic peptide, neurotensin, and somatostatin (Nppb, Nts, and Sst) markers, and a low level of P2X3 in NP3. Histamine receptors (Hth1) were found in NP2 and NP3, and serotonin receptors (Htr1f, Htr2a) were found in NP3 and PEP2. Thus, their data support the existence of at least 3 classes of itch responsive neurones with unique response profiles: lysophosphatidic acid associated with cholestatic disorders may be tuned to NP1 neurones, chloroquine and histamine associated with acute itch may be tuned to NP2 neurones, and mediators, such as IL-31 and cysteine leukotrienes, which are linked to chronic states of inflammatory itch, as well as histamine and serotonin, may engage NP3 neurones (57). These new types and classification of neurones may be closely related to the pathomechanism of dry skin-induced itch.
Keratinocytes
Transient receptor potential cation channel subfamily V member 4 (TRPV4) was reported to be involved in acute itch elicited by exogenously applied histamine and 5-hydroxytryptamine (5-HT) (58, 59). Luo et al. (60) revealed that TRPV4 is selectively expressed by epidermal keratinocytes in mice. Lineage-specific deletion of TRPV4 in keratinocytes reduced itch in AEW-treated mice. Moreover, TRPV4-dependent chronic itch requires 5-HT signalling secondary to activation of distinct 5-HT receptors in AEW as downstream signalling.
AD and allergic contact dermatitis (ACD) are cutaneous diseases characterized by dry skin and chronic itch. Previously, we demonstrated that, possibly through PAR-2 activation in keratinocytes, the cytokine thymic stromal lymphopoietin (TSLP) produced by keratinocytes plays an important role in the development of AD (61). Of note, it was further reported that keratinocytes communicate directly with cutaneous sensory neurones via TSLP to promote itch. Wilson et al. identified the ORAI1/NFAT calcium signalling pathway as an essential regulator of TSLP release from keratinocytes, and TSLP acts directly on a subset of TRPA1-positive sensory neurones to trigger robust itch behaviours (62).
Liu et al. (63) also reported that IL-33 produced and released by keratinocytes is a key cytokine up-regulated in the skin of urushiol-challenged ACD model mice. In this study, IL-33 and its receptor ST2 (expressed in DRG neurones, which innervate the skin) were functionally present in primary sensory neurones and found to lead to pruritus in this model. Studies revealed that hypo-osmotic stress to keratinocytes, such as that noted in AD, and trauma to the skin, such as tape-stripping, promote IL-33 production from keratinocytes (64, 65). Considering the close relationship between itchy-dry skin and AD or ACD, it is highly possible that TSLP and IL-33 produced by keratinocytes play an essential role in the mechanism of dry skin itself.
Spinal cord
Gastrin-releasing peptide system. The GRP and its receptor (GRPR), a Gαq-protein-coupled receptor (GPCR), were reported as itch-specific signalling molecules and expressed in the spinal cord. Intrathecal GRP acts via GRPR to induce scratching behaviour (41). PI3Kγ, a member of lipid kinases that participate in the intracellular signalling cascade, is activated downstream of GPCRs and is related to itch. In a dry skin model of itch, GRPR blockade or PI3Kγ inhibition by intrathecal or systemic route, attenuated the scratching behaviour, suggesting that GRPR is expressed by the central terminals of DRG nociceptive afferents, which transmit itch via the PI3Kγ pathway. These data suggest that the spinal GRP/GRPR system is partly involved in the induction of itch in dry skin (66).
Chemokines. Chemokines are expressed in the central nervous system, where they regulate its function under both physiological and pathological conditions, including neuronal development, synaptic transmission, and disease-associated neuroinflammation (67). Qu et al. (68) reported that C-X-C motif chemokine ligand 10 (CXCL10) and C-X-C motif chemokine receptor 3 (CXCR3) are increased in the DRG in an ACD model, and CXCL10 directly activates a subset of cutaneous DRG neurones through neuronal CXCR3. Of note, AEW treatment induced the expression of CXCR3 and CXCL10 in the spinal cord, and CXCR3–/– mice had fewer scratching responses than control mice. In addition, AEW-induced astrocyte activation was reduced in CXCR3–/– mice, suggesting that the spinal CXCR3 plays an essential role in the pathogenesis of chronic dry skin-induced itch (69).
Toll-like receptors (TLR). TLR are type I transmembrane proteins that can mediate innate and adaptive immunity via recognition of exogenous and endogenous ligands produced after tissue injury. There is increasing evidence that primary sensory neurones express TLRs, e.g. TLR3 and TLR4 (70, 71), and their important roles, such as spinal cord glial activation in neuropathic pain (72, 73). The AEW mouse model exhibited persistent upregulation of TLR4 mRNA and increased TLR4 expression in GFAP-expressing astrocytes in the spinal dorsal horn. TLR4–/– mice exhibited substantial reductions in scratching and alloknesis, a touch-elicited itch in wild-type mice, after AEW. This model also induced TLR4-dependent astrogliosis (GFAP upregulation) in the spinal cord. Intrathecal injection of astroglial inhibitor L-α-aminoadipate reduced AEW-induced itch and alloknesis. Scratching plays an essential role in spinal astrogliosis because AEW-induced astrogliosis was abrogated by placing collars on the neck to prevent scratching. Intrathecal injection of lipopolysaccharide from Rhodobacter sphaeroides (LPS-RS), a TLR4 antagonist, suppressed AEW-induced itch and alloknesis. These findings suggest that spinal TLR4 signalling is important for spinal astrocyte activation and astrogliosis, which may underlie chronic itch and alloknesis (74).
Tumour necrosis factor-α (TNF-α). Emerging evidence suggests that cytokines and chemokines also serve as key itch mediators and/or modulators (75). TNF-α was reported to play a central role in regulating synaptic plasticity in the spinal cord and chronic pain via its receptors, TNFR1 and/or TNFR2 (76). Dry skin itch induced by AEW was reduced by the administration of thalidomide (TNF-α-synthesis inhibitor) and etanercept (TNF-α antagonist), and in TNFR1/R2 double-knockout mice. AEW treatment induced TNF-α expression in the skin, DRG, and spinal cord, and TNFR1 expression only in the spinal cord. Thus, these findings suggest that TNF-α/TNFR1 signalling is partly required for the full expression of chronic itch in dry skin via peripheral and central mechanisms (77).
Others
Zeta chain-associated protein kinase 70. The T-cell signal pathway was reported to function in dry skin pruritus (78). Zeta chain-associated protein kinase 70 (ZAP70), as a T-cell receptor, may induce IL-2 secretion and promote NGF secretion in skin (79). After AEW treatment, 22-month-old AEW mice exhibited increased spontaneous scratching compared with 5-month-old AEW mice. ZAP70 expression was significantly increased, in addition to the secretion of IL-2 and NGF in 22-month-old AEW mice compared with 5-month-old AEW mice. This study revealed that increased ZAP70 is involved in dry skin in pruritus in elderly people, probably due to increased secretion of IL-2 and NGF (80).
Toll-like receptor 3. TLR3 was found to be an important receptor in murine itch signalling, and is expressed by sensory nerves and DRG in mice. TLR3 is also expressed by mast cells and keratinocytes (81). AEW treatment elicited a marked 25-fold increase in TLR3 expression in the skin, but not in the DRGs. Moreover, AEW treatment induced marked NGF upregulation in the skin, which was TLR3-dependent. Spontaneous itch was eliminated in TLR3–/– mice. Thus, TLR3 and its upregulation in the dry skin are important for the induction and sensitization of chronic itch (82).
Opioids. Previous studies have identified 4 major types of opioid receptors, μ-type (MOR, a receptor for β-endorphins), κ-type (KOR, a receptor for dynorphins), δ-type (a receptor for enkephalins), and nociception (a receptor for nociceptin/orphanin FQ). Activation of μ-opioid receptors is thought to induce pruritus, whereas activation of κ-opioid receptors is believed to have suppressive effects (18, 83). We previously reported that the κ-opioid system was downregulated in the epidermis of patients with AD, and that psoralen-ultraviolet A (PUVA) therapy downregulated the μ-opioid system and restored the κ-opioid system, concomitant with a decrease in the visual analogue scale (VAS) score (84). Spontaneous scratching after AEW treatment was significantly suppressed by subcutaneous injection of μ-opioid antagonists, such as naloxone and naltrexone (26), and the κ-opioid agonist nalfurafine (85), presumably via both peripheral and central mechanisms.
Hyperknesis
The term “hyperknesis” was proposed as an umbrella term encompassing the state in which there is enhanced itch to normally itch-provoking stimuli or lowered itch threshold to a given stimulus (86). The mechanisms of hyperknesis are not clear, and it remains unknown which type of afferents mediate the mild itch resulting from punctate stimuli (87). Hyperknesis may be mediated by type-I Aδ fibres through a central mechanism when secondary to itch provocation or an actively itchy skin lesion (88).
Akiyama et al. (27) reported a significant increase in the number of scratching bouts evoked by intradermal injections of a PAR-2 agonist and 5-HT under dry skin conditions. Moreover, DRG cells from AEW-treated mice exhibited significantly larger responses to the PAR-2 agonist and 5-HT. Furthermore, enhanced responses of lumbar superficial dorsal horn neurones to intradermal PAR-2 agonist in this model have been reported (89). This implies that acute itch elicited by certain pruritogens, such as 5-HT and PAR-2 agonists, is enhanced in chronic itchy skin. This reflects hyperknesis, which is consistent with the sensitization of itch-signalling pathways.
Alloknesis
Innocuous mechanical stimuli were reported to elicit itch when delivered within a region of normal skin surrounding a site of experimental itch induced by the intradermal injection of histamine, a phenomenon known as itchy skin or alloknesis (90). Alloknesis may reflect a central mechanism in which the activation of low-threshold mechanoreceptors excites sensitized itch-signalling neurones in the spinal cord. Innocuous mechanical stimulation elicited scratching when delivered at the edge of a region of AEW treatment in mice, suggesting the presence of alloknesis in this animal model of chronic dry skin itch (29).
Merkel cells, the touch receptors in the skin, were reported to make “synapse-like” contacts with type I slowly adapting afferents (91). Feng et al. (92) reported that alloknesis in dry skin is associated with a loss of Merkel cells. Targeted genetic deletion of Merkel cells and its associated mechanosensitive Piezo2 channels produced alloknesis. Chemogenetic activation of Merkel cells protected against alloknesis in dry skin. These data suggest that cutaneous Piezo2 channel-Merkel cell signalling is critical in modulating the conversion of touch to itch.
Chronic itch is clinically correlated with the development of mood disorders, such as anxiety and depression, predominantly in dermatological patients (7). The psychological burden produced by chronic itch was reported with high incidences of suicidal motivation (21.1%) and psychiatric illnesses (70%) (93). Zhao et al. (94) reported that AEW mice developed anxiety-like symptoms 2–3 weeks and depression-like phenotypes 3–4 weeks after AEW treatment, suggesting that mood impairment due to chronic itch evolves over time. The mood impairment behaviours were significantly related to the itch-associated behaviour. They also demonstrated primary disturbance of the hypothalamic pituitary adrenal (HPA) axis function in AEW-treated mice with chronic itch.
The amygdala is the key brain region for the generation of anxiety (95). Recently, Sanders et al. (96) reported that acute itch stimuli, such as histamine, induced anxiety-like behaviour and increased neurone activity in a subpopulation of the amygdala in adult mice. These results highlight the importance of itch-responsive amygdala neurones in the regulation of itch-related effects and behaviour, which may also apply to chronic itch conditions due to dry skin.
Since the mechanisms of dry skin-induced itch in animal models were reported, there have been many studies on the management of dry skin-induced itch (Fig. 2 and Table I).
Table I. Therapies for dry skin-induced itch
Antihistamines
Second-generation H1-antihistamines (e.g. bepotastine) were reported to be beneficial for pruritus in patients with AD (97). We recently demonstrated that bepotastine downregulated NEFs (NGF and ARTN) mRNA in normal human epidermal keratinocytes. The alteration was mediated by the transcription activity of AP-1- and/or NF-kB-dependent mechanisms via the histamine H1 receptor. These results provide therapeutic evidence that second-generation H1-antihistamines may be effective for controlling itch associated with epidermal nerve density in dry skin conditions (98). Another report found that topical application of H1 (diphenhydramine) and H2 (famotidine)-antihistamines prevented epidermal hyperplasia in mice whose skin barrier was disrupted by acetone treatment (99). Similar to many intractable pruritic conditions, AEW-induced itch is thought to be histamine-independent; however, antihistamines may partly improve skin barrier function and epidermal hyper-innervation in dry skin conditions.
Emollients
In our previous study, immediate and delayed application of emollients, e.g. hydrophilic petrolatum and heparinoid cream, onto acetone-induced dry skin reduced the number of penetrated intraepidermal nerve fibres and NGF levels in the mouse skin (33). In addition, application of gel-like moisturizing lotion (TSG), which contained water, glycerin, urea, methyl paraben, propyl paraben, and agar, reduced the number of infiltrated intraepidermal nerve fibres, and induced higher expression of Sema3A in the epidermis of AEW-treated mice. This suggests that topical application of TSG attenuates itch induced by chronic dry skin through a mechanism involving the inhibition of epidermal hyperinnervation (100).
Phototherapy
UV-based therapies, such as PUVA and narrowband-ultraviolet B (NB-UVB), are efficacious in the treatment of chronic pruritus in patients with AD (101) and psoriasis (102). In our previous study, PUVA therapy reduces epidermal hyperinnervation in patients with AD (103). Furthermore, in the acetone induced-dry skin mice model, PUVA, PUVA+betamethasone valerate ointment (BV), NB-UVB, and excimer lamp treatments significantly reduced the intraepidermal nerve growth induced in this model. PUVA+BV and NB-UVB also normalized the abnormal expression of NGF and Sema3A in the epidermis (104).
In addition, we reported that excimer lamp irradiation of nerve ?bres formed by cultured DRG neurones induced degenerative changes in these ?bres. We demonstrated that attaching a cut-off excimer ?lter (COF) to the lamp, thus decreasing cytotoxic wavelengths, reduced hyperinnervation and the production of cyclobutane pyrimidine dimer, a DNA damage marker, in the acetone-induced dry skin mouse model. This suggests that the antipruritic effects of excimer lamp irradiation with COF are due to the induction of epidermal nerve degeneration and reduced DNA damage (105).
Opioids
Dry skin-related itch in animal models was suppressed by μ-opioid receptor antagonists (26) and κ-opioid receptor agonists (85). Clinically, μ-opioid receptor antagonists and κ-opioid receptor agonists were found to inhibit itch in dry skin-related cutaneous or systemic diseases (106, 107).
Nalbuphine is a synthetic opioid analgesic, a mix of κ-opioid receptors agonist-μ-opioid receptors antagonist, clinically indicated for moderate to severe pain (108). A systematic review suggested that nalbuphine is a superior treatment option for opioid-induced pruritus because of its antagonistic effects and high affinity to the μ-opioid receptors (109).
Neurotropin
Neurotropin, a non-protein extract isolated from the inflamed skin of rabbits inoculated with vaccinia virus, was reported as an effective treatment for antihistamine-resistant pruritus in a multicentre, open-label, small sample study (110). We found that neurotropin inhibits NGF-induced neurite outgrowth of DRG neurones in vitro (111). Moreover, the intraepidermal nerve density in acetone-treated mice was reduced by the intraperitoneal administration of neurotropin, probably through the expression of Sema3A in the epidermis (112).
Phospholipids
Eicosapentaenoic acid (EPA, 20: 5n-3) and docosahexaenoic acid (DHA, 22: 6n-3) are representative omega 3 (n-3) polyunsaturated fatty acids (PUFA). Previous studies suggested that n-3 PUFA and related monohydroxy metabolites play an essential role in skin homoeostasis because their content within the skin regulates the skin barrier function (113, 114). Supplementation of fish oil, a well-known source of n-3 PUFA, in an acetone-induced dry skin rat model restored the skin barrier defects and improved scratching behaviour (115).
Dietary milk-derived phospholipids (MPLs) and milk-derived sphingomyelin have been reported to have beneficial effects on epidermal functions (116, 117), such as increased SC hydration in normal hairless HR-1 mice (116), and improved skin barrier function in the HR-AD mouse model (117). Recently, we reported that dietary MPLs attenuate the penetration of nerve fibres into the epidermis by reducing epidermal NGF levels and increasing the Sema3A level in a mouse model of acetone-induced dry skin. Thus, dietary MPLs may have beneficial effects for the prevention and/or alleviation of dry skin-induced itch by reducing intraepidermal nerve fibre density (118).
Collagen tripeptide
Collagen tripeptide is a highly purified, non-antigenic, low allergenic collagen fraction that is known to have many biological effects, such as enhancing hyaluronic acid production in human dermal fibroblasts in vitro and in murine skin in vivo (119). Oral administration of collagen tripeptide to acetone-induced dry skin model mice improves dry skin and normalizes axon-guidance factors in the epidermis, in addition to reducing pruritus (120).
Antioxidants
Oxidative stress has long been proposed to play a role in the pathogenesis of itch-related skin and systemic diseases, including AD, psoriasis, and chronic renal failure (121). Oxidants were demonstrated to induce histamine-independent itch via the activation of TRPA1 in mice (122). Zhou et al. (123) reported that antioxidants were systematically effective in reducing the scratching bouts of AEW-treated mice, possibly through the inhibition of oxidative stress in the periphery (affected skin) and suppression of p-ERK activation in the spinal cord. Thus, antioxidants, such as N-acetyl-L-cysteine and N-tert-butyl-a-phenylnitrone, may have therapeutic effects on dry skin-induced itch.
Film dressings
More recently, we reported that the application of film dressings, which are used for wound treatment to provide an appropriately moist environment and act as a barrier to contamination, alleviated the epidermal hyperinnervation and alloknesis in the AEW-induced dry skin model mice. Film dressings may reduce itch hypersensitivity of the skin (124). Consistent with this, we and others found that the level of NGF in the mouse epidermis significantly decreased by occlusion with emollients (33) or a vapour-impermeable membrane (24) after skin barrier disruption by tape-stripping or acetone. This suggests that skin moisturization prevents epidermal hyper-innervation induced by barrier disruption and mechanical stimuli to the skin.
This review presented recent knowledge regarding the mechanisms of dry skin-induced itch and its management. A decline in skin barrier function is thought to be the primary cause of dry skin-induced itch, as observed in the AEW model, the most well-known mouse model of dry skin. Many kinds of mediators, receptors, and channels are involved in itch signalling among the skin nervous system, skin cells, and central nervous systems, including Mrgprs, TLR, cytokines, and TRP channels. Continued studies are required to better understand these complex interactions and to develop antipruritic drugs to improve the quality of life of the patients.
This work was partly supported by Research Fellowship from the Japan Society for the Promotion of Science (18F18410), Strategic Research Foundation Grant-aided Project for Private Universities from MEXT (S1311011), and KAKENHI (18K07396).
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize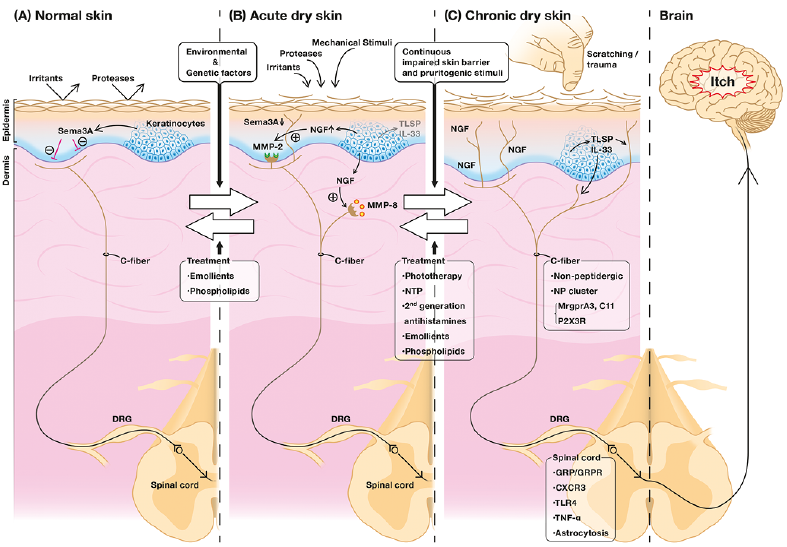 Click to show fullsize
Click to show fullsize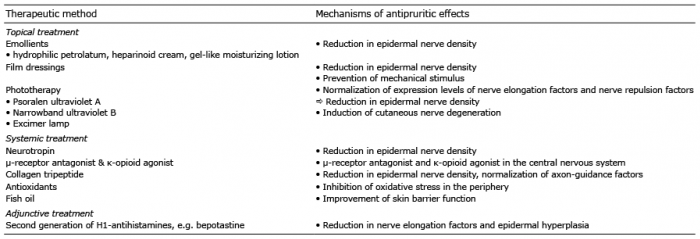 Click to show fullsize
Click to show fullsize