
From the 1Human Performance Laboratory, University of Calgary, Calgary, Alberta, Canada and 2Metabolic Technologies, LLC, Missoula, MT, USA
Objective: To determine the effects of the leucine metabolite β-hydroxy-β-methylbutyrate (HMB) on strength, muscle mass, and contractile material in muscle wasting induced by onabotulinumtoxin type-A (BoNT-A) injection into the quadriceps femoris muscles of New Zealand white rabbits.
Methods: A total of 21, female rabbits were divided into 3 groups (n = 7, each). Group 1 (Control) received intramuscular injection of saline. Groups 2 and 3 received intramuscular injection of BoNT-A (3.5 units/kg), with group 3 receiving supplementation with HMB (120 mg/kg-BW/day). Muscle morphology, mass, and strength were assessed 8 weeks later in both injected and non-injected contralateral limbs.
Results: Injected muscle strength of group 2 (BoNT-A) and group 3 (BoNT-A+HMB) was reduced by 63% and 60%, respectively, compared with Controls (p < 0.0001). Strength in contralateral muscles of group 2 was reduced by 23% vs Controls (p <0.002), while in group 3, strength was similar to Controls. Muscle mass in the injected muscles of the BoNT-A and BoNT-A+HMB groups was significantly reduced, by 46% and 48%, respectively.
Conclusion: While HMB did not prevent loss of muscle strength and mass in the BoNT-A-injected musculature, it prevented significant loss of contractile material in the injected musculature and strength loss in the contralateral non-injected musculature.
Key words: botulinum toxin; Botox®; β-hydroxy-β-methylbutyrate supplementation; muscle strength; muscle mass; skeletal muscle.
Accepted Jul 23, 2021; Epub ahead of print Aug 25, 2021
J Rehabil Med 2021; 53: jrm00229
Correspondence address: John C. Fuller, Jr, Metabolic Technologies LLC 135 West Main St. Suite B Missoula, MT 59802, USA. E-mail: Fuller@mti-hmb.com
Doi: 10.2340/16501977-2866
Onabotulinum toxin A (BoNT-A) is currently approved for cosmetic procedures and for the treatment of many conditions, such as migraine headaches and neuromuscular conditions associated with spasticity, such as cerebral palsy or post-stroke in USA. Once injected into the muscle BoNT-A causes muscle paralysis, which is reversible. The current study examined the local and distant detrimental effects of BoNT-A on decreasing muscle wasting and muscle function in a rabbit model, and the effects of dietary use of HMB (β-hydroxy-β-methylbutyrate), a leucine metabolite, on these detrimental effects. BoNT-A was injected into 1 leg of a rabbit model and muscle mass and strength measured in both the injected and non-injected legs. BoNT-A caused a loss of muscle mass in the injected legs. It also caused a loss of strength in the injected legs and a somewhat reduced loss of strength in the non-injected legs. This suggests that BoNT-A injections can cause a decrease in function in distant, non-target muscles. HMB partially reversed some of the detrimental side-effects of BoNT-A, being shown to be effective in maintaining muscle strength in the non-injected leg.
Muscle mass is maintained by a balance between protein synthesis and degradation. However, when this balance is disturbed by either an increase in protein breakdown or a reduction in protein synthesis, muscle wasting occurs. Muscle wasting has significant clinical consequences, including muscle weakness, fatigue, morbidity, and it is an important contributing factor for reduced quality of life (QoL). Treatment modalities to prevent muscle wasting include exercise, inhibitors of inflammatory cytokines, and anti-inflammatory cytokines (1). Recently, there has been an increase in dietary supplement use aimed at increasing muscle mass, or preventing muscle atrophy in healthy, active, and sedentary people, in high-performance athletes, and in patients exposed to prolonged bed rest (2–5). However, the efficacy of dietary supplements in preventing strength loss and atrophy is a matter of debate (3).
Onabotulinumtoxin type-A (BoNT-A) is a potent neurotoxins (6, 7). Once injected into the target muscle, BoNT-A binds with high affinity to the neuromuscular junction, preventing acetylcholine release, and thereby inducing dose-dependent muscle paralysis (8, 9). Due to its extreme potency and specificity, BoNT-A has been used for both therapeutic and scientific interventions (10–12).
Localized BoNT-A injections are used clinically in an increasing number of neuromuscular disorders, with the primary aim of relaxing the hyperexcitability of peripheral nerve terminals; for example, in patients with cerebral palsy or following a stroke (11, 13). In experimental settings BoNT-A has been used to induce muscle weakness in an attempt to mimic muscle atrophy following injury, denervation, in elderly patients (14–16), or to determine the effects of muscle weakness on bone and joint health (17, 18). BoNT-A-induced muscle weakness can also be used to investigate strategies for the prevention of, or the reverting of, strength loss and muscle atrophy (19).
β-Hydroxy-β-methylbutyrate (HMB) is naturally produced from the amino acid leucine (20). As a dietary supplement, HMB has been used to improve strength and gain lean body mass, reduce muscle protein degradation, and improve recovery following exercise in trained, untrained, and elderly people (5, 21–24). Supplemental HMB has also been shown to increase lean body mass in elderly subjects without exercise (2) and to retain lean body mass during bed rest (4). Other studies have shown HMB can improve protein turnover in muscle atrophy conditions, such as AIDS (25), trauma (26) and cancer (27), and in endotoxin-induced septic rats (28).
Despite the increasing use of HMB as a dietary supplement, there have been no definitive studies showing the efficacy of HMB to prevent muscle weakness under conditions known to produce muscle paralysis and consequent atrophy.
The objective of the current study was to evaluate the effects of supplemental HMB on strength, muscle mass, and contractile material following BoNT-A-induced weakness in the quadriceps femoris of New Zealand white (NZW) rabbits. It was hypothesized that HMB supplementation would reduce the loss of strength and atrophy in both BoNT-A-injected target muscles and non-injected, non-target muscles.
Experimental design
A total of 21 female NZW rabbits (43 weeks old) (Covance Research Products, Inc., Greenfield, IN, USA) were used in this study. All procedures were approved by the Animal Care Committee of the University of Calgary. Rabbits were allowed normal activity in a 65 × 45 × 30 cm cage and had ad libitum access to food and water.
The rabbits were divided into 3 groups:
BoNT-A injection protocol
The rabbits were injected with Clostridium botulinum type-A neurotoxin complex (Onabotulinum Toxin A, Allergan, Inc., Toronto, Ontario, Canada), reconstituted with 0.9% sodium chloride to 20 U/ml. The rabbits received intramuscular BoNT-A injections at a total dose of 3.5 U/kg. Injections were randomized to either the right or left quadriceps. The anterior compartment of the thigh was isolated by manual palpation and the quadriceps was visually divided into superior and inferior halves. Each half was subdivided into a medial, central, and lateral section. One sixth of the total BoNT-A dose was injected into each section to increase its diffusion and to equally distribute the toxin throughout the different portions of the quadriceps musculature (10, 12).
The Control group received randomized intramuscular saline injections. The total volume of injected saline was the same as the total volume of BoNT-A injections into the experimental group animals. The other 2 groups of rabbits received a single intramuscular BoNT-A injection, as described above. All treatment groups were evaluated 8 weeks after the injections.
Feeding and supplementation
The food provided for the Control group and BoNT-A group was a high-fibre diet (Laboratory Rabbit Diet HF 5326, LabDiet, Richmond, IN, USA), while the BoNT-A+HMB group rabbits received the same basal diet, custom formulated with 0.44% calcium β-hydroxy-β-methylbutyrate monohydrate (CaHMB, Metabolic Technologies, Inc. (MTI), Ames, IA, USA). The CaHMB used in the study was determined to be greater than 99% pure, and the HMB in the rabbit food formulation was determined to be 88.1% of expected and both the purity and formulation were within acceptable limits (MTI). The formulation for the food was calculated such that an estimated daily dosage of 120 mg/kg body weight/day would be eaten by each of the rabbits on the HMB treatment. This dosage is equivalent to the human dosage of 38 mg/kg body weight/day when corrected for body surface area (29).
Determination of food intake, knee extensor strength, muscle mass, and contractile material
The food intake was measured daily throughout the experimental period by weighing the individual food hoppers across the groups at the same time of the day, using a commercial scale with a resolution of 0.5 g.
The primary outcome measures were the isometric knee extensor torque, measured via femoral nerve stimulation, the mass of the individual quadriceps muscles, and the percentage of contractile material in the injected and contralateral non-injected musculature. Isometric knee extensor strength was measured in the injected and contralateral musculature 8 weeks after the BoNT-A injection. Following nerve cuff implantation, rabbits were secured in a stereotactic frame using bone pins at the pelvis and femoral condyles. Isometric knee extensor forces at 80° of knee flexion were measured using a strain-gauged, calibrated bar placed over the distal portion of the rabbit’s tibia (10).
Stimulation of the knee extensor musculature (Grass S8800 stimulator; Astro-Med Inc., Longueil, Quebec, Canada) was performed at a voltage 3 times higher than the alpha motoneurone threshold, to ensure activation of all motor units (30). Stimulation duration was 500 ms, pulse duration 0.1 ms, and frequency of stimulation 200 Hz.
Following knee extensor strength assessment, animals were sacrificed by an overdose of Euthanyl (MTC Pharmaceutical; Cambridge, Ontario, Canada) via heart puncture. Wet mass for the individual quadriceps femoris muscles was determined using a commercial scale with a resolution of 0.01 g.
The percentage of contractile material was determined histologically. The central third of the quadriceps muscle was embedded in paraffin (automatic paraffin processor, Leica TP 1020) and cut cross-sectionally with a microtome (Leica RM 2165). For every 100 µm, an 8 µm section was collected for staining with haematoxylin-eosin (H&E) (Leica ST5010) (19, 31, 32). Photographs were taken from each stained section using an Axionstar plus microscope (Carl Zeiss AG) with a 5× magnification objective. A customized Matlab program (MatLab, R2019a) was used to calculate the percentage of contractile material for at least 50% of the total cross-sectional area of each muscle (33).
Data analysis
The data were analysed using Proc GLM in SAS (SAS for Windows 9.4, SAS Institute, Inc., Cary, NC). A 1-way analysis of variance (ANOVA) model was used with the main effect of treatment. For actual muscle weights and strength measurements, the body weight of the rabbit was used as a covariate. Means reported are least squares means with the standard error of the mean. The p-values given for overall treatment are from the main effect model, while individual means were compared using the least square means predicted difference. A post hoc χ2 analysis was performed on the histology measurement of the contractile material between the BoNT-A alone and the BoNT-A+ HMB groups. A p-value ≤ 0.05 was determined to indicate significance.
Body weight and food intake
The Control, BoNT-A, and BoNT-A+HMB group rabbits weighed 4.4 ± 0.1, 4.1 ± 0.1, and 4.4 ± 0.3 kg at the start of the study, and 4.2 ± 0.1, 4.0 ± 0.1, and 4.0 ± 0.1 kg at the end of the 8-week study period, respectively. There were no differences in body weights across groups either before or after the control or BoNT-A treatments. Food intake over the 8-week study was 169 ± 8, 136 ± 8, and 129 ± 8 g/day for the control, BoNT-A, and BoNT-A+HMB rabbits, respectively. The BoNT-A and BoNT-A+HMB groups ate significantly less food due to decreased food intake after the BoNT-A injections (p<0.005). Mean food consumption was reduced 52% in the BoNT-A group and 35% in the BoNT-A+HMB group during weeks 2 and 3 following the injection. During the 2-week period immediately following the BoNT-A injection, a mean HMB dosage received by the BoNT-A+HMB group was 80 ± 17 and 86 ± 22 mg/kg body weight/day during weeks 2 and 3, respectively. During this period, the rabbits consumed approximately two-thirds of the target dosage; however, over the 8-week study period, a mean dosage was 138 ± 10 mg/kg body weight/day, or approximately 15% above the target dosage.
Muscle strength
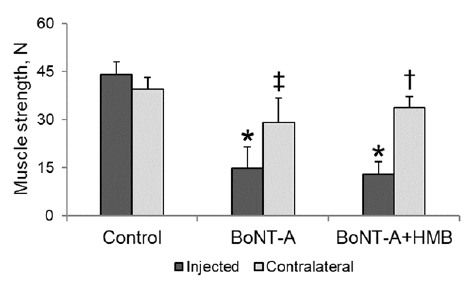
There was no difference in strength between the saline-injected and contralateral musculature for the Control group rabbits. Strength in the injected musculature was 44 ± 2, 16 ± 2, and 18 ± 2 N for the control, BoNT-A, and BoNT-A+HMB rabbits, respectively. Strength of the BoNT-A injected muscles was significantly (p < 0.0001) reduced, by 63% and 60%, in the BoNT-A and the BoNT-A+HMB group rabbits, respectively, compared with the Control group rabbits. Supplementation with HMB did not prevent strength loss in the BoNT-A+HMB group rabbits compared with the BoNT-A group rabbits (Fig. 1).
Strength in the contralateral musculature was 45 ± 2, 34 ± 2, and 41 ± 2 N for the Control, BoNT-A, and BoNT-A+HMB group rabbits, respectively. Strength was significantly (p < 0.002) reduced, by 23%, in the contralateral musculature of BoNT-A group rabbits compared with Control group rabbits. Strength was significantly greater (p<0.04) in the contralateral musculature of BoNT-A+HMB (41 ± 2 N) than in the BoNT-A group rabbits and was not significantly different from strength in the Control group rabbits (Fig. 1).
Fig. 1. Mean muscle strength ( ± standard error of the mean; SEM) in rabbit quadriceps musculature for the control (saline-injected), onabotulinumtoxin-A (BoNT-A), and BoNT-A+HMB group rabbits. Results for the injected and contralateral muscle. Groups are shown. (*p < 0.0001 compared with Control group rabbits injected leg strength, †p < 0.04 compared with BoNT-A group rabbits contralateral leg strength, ‡p < 0.002 compared with Control group contralateral leg strength).
Muscle mass
There was no difference in muscle mass for the saline-injected and contralateral muscles of the Control group rabbits. Muscle mass in the injected musculature was 33 ± 3, 17 ± 2, and 17 ± 3 g for the control, BoNT-A, and BoNT-A+HMB group rabbits, respectively. Muscle mass in the injected limbs was significantly (p < 0.0001) reduced, by 46% and 48%, for the BoNT-A and BoNT-A+HMB groups, respectively, compared with the Control group rabbits. Supplementation with HMB did not prevent muscle mass loss in the injected musculature of the BoNT-A+HMB group compared with the BoNT-A group rabbits (Fig. 2).
Fig 2. Mean muscle mass ( ± standard error of the mean; SEM) of rabbit quadriceps musculature for the control (saline-injected), onabotulinumtoxin-A (BoNT-A). and BoNT-A+HMB rabbits. (*p < 0.0001 compared with Control group rabbits).
Muscle mass values in the contralateral non-injected musculature were 33 ± 2, 32 ± 2, and 34 ± 3 g for the control, BoNT-A, and BoNT-A+HMB group rabbits, respectively, and remained similar across all 3 experimental groups.
Contractile material
There was no difference in the percentage of contractile material for the saline-injected and contralateral musculature of the Control group rabbits. The amount (area) of contractile material was measured in the injected and contralateral legs of all animals, and the contractile material for the injected musculature was 93 ± 5, 75 ± 16, and 85 ± 4% for the control, BoNT-A, and BoNT-A+HMB group rabbits, respectively. Following the BoNT-A injection, the contractile material was significantly (p < 0.006) reduced, by 19% and 8%, in the BoNT-A and BoNT-A+HMB group rabbits. Least square means analysis showed that with the HMB diet intervention the decrease contractile material was not significant compared with the Control group. In addition, a post hoc χ2 analysis between the BoNT-A and BoNT-A+HMB groups showed that HMB-fed rabbits had a significantly greater amount of contractile material compared with the BoNT-A group animals (p < 0.005; Fig 3).
There were no differences in the percentage of contractile material in the contralateral non-injected musculature, which were 90 ± 2%, 87 ± 7%, and 92 ± 4% for the control, BoNT-A, and BoNT-A+HMB group rabbits, respectively (Fig. 3).
Fig. 3. Exemplar histological cross-sectional images showing the percentage of contractile material (haematoxylin and eosin (H&E): red staining) and non-contractile material (white colour – primarily fat and connective tissue). The percentage of contractile material for the saline-injected rabbits was 93 ± 5% (top left). Following onabotulinumtoxin-A (BoNT-A) injection, there was a significant reduction in contractile material to 75 ± 16% (middle left) and 85 ± 4% (bottom left) for the BoNT-A and BoNT-A+HMB group rabbits, respectively. There was no difference in the percentage of contractile material for the contralateral non-injected musculature across groups. Black bars indicate 100 μM in the panels
This study showed that HMB supplementation did not prevent muscle mass and strength loss in BoNT-A injected quadriceps femoris muscles of New Zealand white rabbits as originally hypothesized. However, HMB supplementation did help prevent loss of contractile material in the injected muscle, as surmised, and prevented strength loss in the contralateral non-target musculature of rabbits receiving BoNT-A injections.
BoNT-A is a potent neurotoxin that prevents muscle contraction in a dose-dependent manner (8, 34). BoNT-A can produce substantial muscle weakness lasting up to 6 months following injection (33). Therefore, it might be that the high affinity of the toxin with motoneurons reduced the effect of HMB in inducing muscle hypertrophy due to a lack of muscle activation. Also, muscle strength in the BoNT-A and BoNT-A+HMB group rabbits might have been dominated by the inhibitory effects of BoNT-A at 8 weeks post-injection and might have masked true potential differences in strength between these 2 experimental groups. Direct muscle stimulation avoids the inhibitory effects of BoNT-A and has been shown to produce significantly greater strength values in BoNT-A injected muscles than stimulation of the corresponding nerve. Therefore, it cannot be excluded that HMB supplementation might have prevented strength loss in the BoNT-A injected muscles, but that this effect was not observed because of the remnant inhibition of acetylcholine at the nerve endings.
In humans, HMB supplementation has been shown to enhance muscle recovery, increase lean body mass, and increase muscle strength, power, and aerobic performance (5). HMB supplementation in humans has also been shown to preserve muscle mass over a 10-day period of bedrest, a condition which results in muscle atrophy (4). Muscle mass is determined by the balance of protein synthesis and protein degradation, and HMB has been shown to affect both processes, resulting in a decrease in protein breakdown and an increase in protein synthesis (35, 36). In vitro studies have shown that HMB decreases proteolysis through attenuation of the induction of the ubiquitin-proteasome proteolytic pathway when muscle protein catabolism is stimulated by either proteolysis inducing factor (PIF), lipopolysaccharide (LPS), or angiotensin II (37). In an animal model of cancer-induced muscle loss in mice, Smith et al. (36) also demonstrated a decrease in the ubiquitin-proteasome pathway as well as a stimulation of protein synthesis. Still other in vitro studies have demonstrated that HMB also attenuates the activation of caspases-3 and -8 proteases whose activation increases protein degradation through the ubiquitin-proteasome pathway (37, 38). Other in vitro studies showed that HMB stimulates protein synthesis through mammalian target of rapamycin (mTOR) phosphorylation (39), and Wilkinson et al. (40) demonstrated that HMB increases protein synthesis in humans through the mTOR pathway, which was accompanied by an overall decrease in muscle protein breakdown.
However, the current study did not show that HMB prevented loss of muscle mass and strength in the injected musculature. Possible reasons for why no such effect of HMB supplementation was observed are: (i) the HMB dosage was not sufficient to elicit an anabolic response; (ii) the BoNT-A dosage used was too high and the effects of HMB were overshadowed by the BoNT-A effects on the target muscles; (iii) an intact neuromuscular junction is necessary for HMB to have an effect on strength; or (iv) HMB needs to be combined with muscle training.
The dosage of HMB used was calculated using an effective human dosage, and the depressed food intake caused by the BoNT-A injection was not taken into consideration. Thus, the rabbits consumed only approximately two-thirds of the targeted dosage during the second and third weeks after the injections. In exercising humans, HMB has been shown to have a dose response when approximately 19 and 38 mg/kg body weight/day was supplemented (21). However, it is unknown what dosage of HMB might be most effective in the current study rabbit model, and only a single dosage was used in the current study. Kovarik (28) demonstrated deceased protein turnover in endotoxin-induced septic rats, at a dosage of 500 mg/kg body weight/day, which is equivalent to a dosage of approximately 250 mg/kg body weight/day in rabbits or approximately 3 times the dosage given to the rabbits in the current study following the BoNT-A injection. Therefore, an even higher dosage of HMB, or a different dosing method to ensure that the rabbits receive the targeted dosage throughout the study period (i.e. gavage or intraperitoneal injection) may have resulted in a greater effect. Secondly, the dosage of BoNT-A used may have caused a severe inflammatory response, with an exacerbated loss of contractile material that the HMB dosage was unable to overcome (33). While sparing of contractile material in the BoNT-A-injected muscle was observed in the current study with HMB treatment, it is unclear why functional strength improvements were not seen in the injected limb. Perhaps more recovery time would have shown an effect in strength measurement in the injected limbs, as was seen in the contralateral limbs. While HMB can positively affect protein turnover in in-vitro muscle cell cultures exposed to agents that initiate protein loss, the complexity of the animal model used in the current study could demonstrate the importance of continued neuromuscular communication and muscle activity for HMB to exert an effect on maintaining muscle mass. Therefore, the effect of BoNT-A on muscle innervation could have contributed to the observation of no improvement in strength in the injected leg with HMB, despite the observation of contractile material sparing. Future studies should examine different BoNT-A dosages to identify which dosage HMB might overcome the BoNT-A affect and elicit an anabolic response following injection.
BoNT-A also affects the non-injected, non-target muscles causing atrophy, loss of strength, and loss of contractile material (31). Strength in the contralateral muscles was decreased for the BoNT-A, but not the BoNT-A+HMB group rabbits, while muscle mass was the same for the Control group and BoNT-A-injected group rabbits. These results suggest that either the inhibitory effect of BoNT-A in the contralateral limbs was eliminated in the BoNT-A+HMB group rabbits, but not the BoNT-A group rabbits, or that HMB supplementation preserved the structural integrity of the contralateral muscles. The contralateral muscles in this BoNT-A model can manifest loss of contractile material, which affects strength but not muscle mass, that supplementation with HMB might have prevented, and thus mass and strength were preserved in the BoNT-A+HMB group rabbits. If proven correct, HMB supplementation might be an important therapeutic agent for people with muscle spasticity (cerebral palsy, post stroke) who receive regular BoNT-A treatments. Preservation of structure and strength of non-target muscles in such patients might preserve function in these muscles beyond what is possible with traditional intervention therapies.
HMB supplementation did not prevent loss of muscle strength and mass in the BoNT-A-injected rabbit musculature when tested using femoral nerve stimulation. This result needs to be confirmed using direct muscle stimulation to avoid possible confounding effects associated with BoNT-A inhibition. Furthermore, HMB prevented loss of contractile material in the injected rabbit musculature and loss of strength in the contralateral non-injected muscles suggesting that HMB either decreased BoNT-A-induced inhibition of the contralateral, non-injected muscles, or preserved the structural integrity in non-injected muscles of BoNT-A treated animals. Maintenance of contralateral strength could be important after an immobilizing injury, and further study is necessary.
The authors thank the Canada Research Chair Programme, The Killam Foundation, and The Canadian Institutes of Health Research. In addition, the authors wish to acknowledge Metabolic Technologies, LLC, Missoula, MT, USA for funding this study.
Conflicts of interest. John C. Fuller Jr. works for Metabolic Technologies, LLC who funded the study and also sell HMB. All other authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize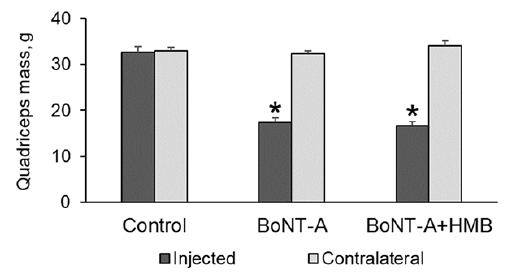 Click to show fullsize
Click to show fullsize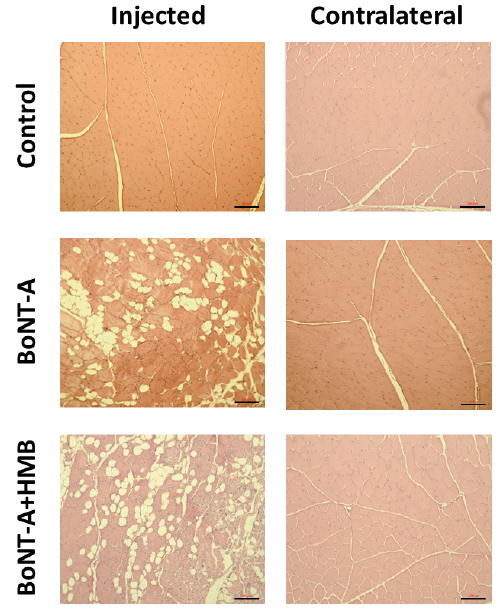 Click to show fullsize
Click to show fullsize