


Gerd G. Gauglitz1*, Helene Callenberg1*, Günther Weindl2 and Hans C. Korting1
1Department of Dermatology and Allergology, Ludwig-Maximilian-University, Munich and 2Institute of Pharmacy, Free University of Berlin, Berlin, Germany *Both authors contributed equally to this manuscript and should be considered as first authors.
Recognition of Candida albicans is mediated by several classes of pattern-recognition receptors, including Toll-like receptors and C-type lectin receptors. Cell wall components of C. albicans, interact with the pattern-recognition receptors, which are expressed by different cells, primarily antigen-presenting cells. This review aims to discuss the different pattern-recognition receptors responsible for recognition of special structures of C. albicans, which are known to activate intracellular signals that finally lead to directed and efficient host defence. Key words: Candida albicans; pattern-recognition receptor; pathogen-associated molecular pattern; C-type lectin receptor; toll-like receptor.
Accepted August 15, 2011.
Acta Derm Venereol 2012; 92: 291–298.
Hans C. Korting, Department of Dermatology and Allergology, Ludwig-Maximilian-University, Frauenlobstrasse 9–11, DE-80337 Munich, Germany. E-mail: HansChristian.Korting@med.uni-muenchen.de
Candida species are common fungal pathogens in humans. The spectrum of clinical diseases caused by Candida spp. encompasses various types of manifestations, including localized mucocutaneous infections and systemic deep-seated infections of various internal organs (1–4) representing septicaemia. The polymorphic yeast Candida albicans can coexist as a commensal within the microbial flora of the human being on mucosal surfaces. Under physiological conditions, C. albicans may thrive harmlessly inside its accommodating host. In case of disturbed equilibrium of the microbial flora and/or additional immunosuppression of the host, superficial infection of the skin or the bordering mucosal surfaces, or invasive candidiasis, where C. albicans disseminates throughout the blood and infects remote inner organs may occur (5). The frequency of mucosal and cutaneous fungal infections is increasing worldwide, with oral candidiasis being the most common human fungal infection (6). Various clinical presentations of oral candidiasis include acute pseudomembranous candidiasis, chronic atrophic candidiasis, perlèche, candidal cheilitis, candidal leukoplakia, and chronic mucocutaneous candidiasis (6). Disseminated candidiasis represents one of the most frequent types of nosocomial invasive infections today, with a lethality rate of up to 30–40% (7). Given the great levels of morbidity and mortality associated with nosocomial candidiasis, the pathogenic adaptation of C. albicans has been the topic of extensive investigations (8).
In order to be capable of inducing such a variety of infections C. albicans can survive in several anatomically distinct sites and expresses several virulence factors. The phenomenon of phenotypic switching from yeast- to filament-growth is just one, but crucial, factor that contributes to the virulence of C. albicans. It provides a basis for activating different receptors leading to different immune responses. Other virulence factors of C. albicans include adhesion factors, thigmotropism (ability to identify intercellular junctions at the mucosal surface by contact sensing and their targeted penetration), and secretion of various hydrolytic enzymes, such as lipase, phospholipase, and proteinase (reviewed in 9, 10).
The interaction between virulence factors of C. albicans and host defence mechanisms plays a crucial role in determining whether colonization remains harmless or leads to infection of the epithelium and possibly systemic infection (11).
The fungal cell wall contains a glucan and chitin skeleton and a matrix of glycosylated proteins. In C. albicans the major proteins of the cell wall are glycosylphosphatidylinositol (GPI)-anchor-dependent cell wall proteins (GPI-CWPs) with a particular affinity to the skeleton-parts ß-(1,3)-glucan and chitin. CWPs are glycosylated with mannose-containing polysaccharides (mannan), which represent one crucial structure for binding pattern-recognition receptors (PRRs) (12).
Progress in our understanding of the mechanisms responsible for C. albicans recognition, as described below, allows us to suggest an integrated view of how C. albicans is recognized and of how the host innate immune response is activated during candidiasis. Although we are still in the early stages of understanding the complexity of fungal recognition, there are several widely accepted principles that characterize recognition of C. albicans (12).
First, PRRs are expressed by cells of the innate immune system and specific immunity (Table I) to recognize pathogen-associated molecular patterns (PAMPs), which are structural components characteristic for microbial pathogens (Table II). Recognition of PAMPs leads to microbial uptake and, consecutively, to a modulation of gene expression to initiate an adaptive and target-oriented immune response.
Table I. Cell populations and pattern-recognition receptors (PRRs) involved in C. albicans recognition. The main cell populations involved in recognition of C. albicans during the innate immune response include monocytes, macrophages and neutrophils. Dendritic cells are crucial for processing of and antigen presentation to T cells, and therefore to activation of specific immunity. The differential expression of PRRs by these cells is elucidated (extracted from (12))
|
PRRs |
Monocyte |
Macrophage |
Neutrophil |
Dendritic cell |
CD4+ T cell |
|
TLR2 TLR4 TLR6 TLR9 MR Dectin-1 Dectin-2 DC-SIGN CR3 FcγR Galectin-3 |
× × |
× × × × × × × × × × × |
× ×
×
× × |
× × × × × × × × |
× × × |
MR: mannose receptor; TLR: toll-like receptor; DC-SIGN: dendritic cell-specific intercellular adhesion molecule-3-grabbing non-integrin; CR3: complement receptor 3.
Table II. Pattern-recognition receptors (PRRs) sensing Candida-associated molecular patterns. Candida pathogen-associated molecular patterns (PAMPs) are recognized by specific PRRs from three major families: toll-like receptors (TLRs), C-type lectin receptors (CLRs) and nucleotide-binding domain, leucine-rich-repeat-containing receptors (NLRs)
|
PRRs |
Candida PAMP |
Refs |
|
Toll-like receptors |
||
|
TLR2 |
Phospholipomannan |
(29) |
|
TLR4 |
Mannan O-linked mannosil residues |
(18) (20) |
|
TLR9 |
CpG-oligodeoxynucleotides |
(48) |
|
C-type lectin receptors |
||
|
Mannose receptor |
Mannan N-linked mannosil residues |
(111, 112) (20) |
|
Dectin-1 |
β-1,3-Glucan |
(113) |
|
Dectin-2 |
High-mannose structures (Man9GlcNAc2) |
(114) |
|
DC-SIGN |
High-mannose structures |
(61) |
|
Mincle |
Unknown |
(76) |
|
Galectin-3 |
β-1,2-Mannosides |
(81) |
|
SCARF1/CD36 |
β-1,3-Glucan |
(115) |
|
NLRs |
||
|
NLRP3 |
Unknown |
(108) |
Reprinted from (40) with permission from Elsevier.
To date, three main groups of specific receptor systems dubbed PRRs have been distinguished: toll-like receptors (TLRs), C-type lectin receptors (CLRs) and nucleotide-binding domain, leucine-rich repeat containing (NLR) protein family (Table II). As summarized in Fig. 1, each ligand–receptor system activates specific intracellular signalling pathways, which in turn leads to modulation of the various components of the host immune response (12). Also, it is critical to mention that the fully integrated response to a specific pathogen depends on the mosaic of PRRs and receptor complexes that are engaged (12). Importantly, co-stimulation via multiple PAMP–PRR interactions may increase both the sensitivity and the specificity of the immune recognition process (12). Also, members within one family of PRRs can trigger opposite signalling effects, indicating that the ultimate outcome of a pathogen-induced immune response depends on the pathogen signature and the PRRs involved.
TOLL-LIKE-RECEPTORS
The discovery of TLRs in 1987 and their first description in 1997 have significantly contributed to the understanding of the complex mechanism underlying the recognition of microorganisms by the innate immune system. Ever since, many studies have been undertaken that reflect the complexity of this interaction. To date, it has become increasingly clear that recognition of C. albicans by myeloid cells and the resultant signalling leads to a pro-inflammatory response towards the yeast, which has been described as necessary for acquiring resistance to infection (8). Importantly, Moyes et al. (13) showed that epithelial responses appear to be targeted predominantly to the hyphal form with the yeast being non-stimulatory. Notably, to date it is accepted that both TLR-2 and TLR-4 represent the main TLRs involved in the signalling cascades induced by C. albicans, even though, depending on the model utilized, conflicting results have been obtained (14–22).
TLRs are a family of evolutionarily conserved receptors that react to bacterial, viral, or fungal antigens or to endogenous factors released during cell injury (11). TLRs are type-1 transmembrane proteins that are characterized by extracellular leucine-rich repeats (LRRs), followed by a single transmembrane domain and a globular cytoplasmic domain called the toll-interleukin 1 receptor (TIR) domain. Thirteen TLRs have been identified in mammals so far and ten are functional in humans. TLR1, TLR2, TLR4, TLR5 and TLR6 are cell-membrane-associated, whereas TLR3, TLR7, TLR 8 and TLR 9 are localized to the endosomal or lysosomal compartment.
The activation of the TLR signalling pathways originates from the cytoplasmic (TIR) domain that associates with TIR domain-containing adaptors and induces transcription factors, such as nuclear factor (NF)-κB and interferon regulatory factors 3/7 (IRF3/7), followed by enhanced transcription of antimicrobial peptides, cytokines and chemokines that affect innate and adaptive immune responses (23–26) (Fig. 1). Ligation of TLRs on dendritic cells (DCs) also induces expression of co-stimulatory molecules (maturation), and this, in combination with specific cytokine profiles, drives the differentiation of naïve CD4+ T cells (Fig. 1). Several TLRs are involved in inflammatory responses induced by C. albicans, of which TLR2 and TLR4 are the most studied (12). TLR2 and TLR4 are expressed by different cell types of the innate immune system including monocytes, macrophages, dendritic cells, neutrophils, CD4+ cells (27) and epithelial cells (11, 28). Activation of TLR2 signal pathways in these antigen-presenting cells (APCs) by ligation of C. albicans cell-wall components such as phospholipomannan leads to production of cytokines, such as tumour necrosis factor (TNF), interleukin (IL)-1β and IL-10 (29–31), while IL-12 and IFNγ are not induced, resulting in a balance favouring a Th2 cellular response (32). In 2002, Netea et al. (30) demonstrated that blocking TLR2 with a TLR2-specific antibody after stimulation of monocytes by C. albicans resulted in diminished release of TNF and IL-1ß. Also, TLR2−/− mice have been shown to be more resistant to lethal disseminated candidiasis, indicating a TLR2-mediated inhibitory effect on inflammation in this model (33, 34). However, another group has reported a higher susceptibility for disseminated candidiasis in TLR2−/− mice (35). These discrepancies are probably based on different mouse models utilized in these studies and different Candida strains. The immunosuppressive effect induced by TLR2 may indeed explain the increased resistance of TRL2–/– mice to disseminated candidiasis and was found to be mediated through the generation of T-regulatory cells with immunosuppressive potential (33, 34). Interestingly, Ali et al. (36) reported that C. albicans utilize TLR2 to evade the immune system of the host through its hyphae form, while Roeder et al. (37) showed that TLR2 induces the transcription factors NF-κB and c-Jun/activator protein-1 in macrophages in the presence of anti-mycotic-treated C. albicans.
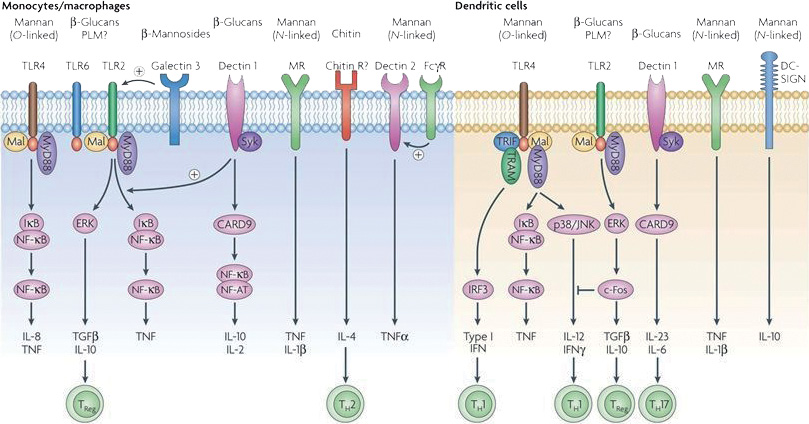
Fig. 1. “The major pattern-recognition receptors (PRRs) involved in the recognition of specific C. albicans pathogen-associated molecular patterns (PAMPs). Stimulation of host response by C. albicans at the cell membrane is mediated by toll-like receptors (TLRs) and C-type lectin receptors (CLRs). TLR4 induces mainly pro-inflammatory signals in monocytic cell types (monocytes, macrophages and dendritic cells (DCs)) through the MyD88–Mal-mediated NF-κB and mitogen-activated protein kinase (MAPK) pathways, while stimulating TH1 responses through IRF3-dependent mechanisms mainly in plasmacytoid DCs. TLR2 stimulates the production of moderate amounts of pro-inflammatory cytokines and strong IL-10 and TGFβ responses. The pro-inflammatory effects of TLR2 can be amplified by dectin-1 and galectin-3, the latter especially in macrophages. In addition to amplifying the effects of TLR2, the non-classical lectin-like receptor dectin-1 induces IL-2, IL-10 and TH17 responses through a Syk–CARD9 cascade, independently of its interaction with TLR2. The classical lectin-like receptor, the mannose receptor (MR), induces pro-inflammatory effects in monocytes and macrophages, whereas chitin-dependent stimulation of these cells induces mainly TH2 responses, although this effect has yet to be demonstrated for C. albicans, and the identity of the chitin receptor is unknown. Other less well characterized pathways include stimulation of tumour necrosis factor (TNF) and IL-1Ra by dectin-2, while engagement of DC-SIGN in DCs induces production of the immunosuppressive cytokine IL-10.” (Reprinted from (12), with permission from Nature Publishing Group).
A partial role for TLR1 and TLR6 in the in vitro recognition of C. albicans and the induction of cytokines by macrophages has been reported, but the absence of these receptors from genetically manipulated animals did not result in an increased susceptibility to infection (38).
TLR4 represents one of the most in-depth studied PRRs, due to its role as the main receptor of bacterial lipopolysaccharides (39). TLR4 recognizes mannans from Saccharomyces cerevisiae and C. albicans (18). In vitro, short linear O-bound mannans of C. albicans are recognized by TLR4 and persuade pro-inflammatory cytokine production, such as TNF (20). However, the role of TLR4 for host defence against invasive candidiasis has been hotly debated, due to conflicting results from different laboratories (40).
In the first experimental model of disseminated C. albicans infection, Netea et al. (30) reported an increased susceptibility of TLR4-defective mice to disseminated candidiasis due to a decreased induction of KC and MIP2. This results in a deficient recruitment of neutrophils and higher fungal burden in the kidneys of TLR4−/− mice compared with that of WT mice (31). The human TLR4 polymorphisms Asp299Gly and Thr399Ile led to a higher susceptibility for Candida bloodstream infections (41). In addition, stimulation of DCs by C. albicans induces TLR4-dependent production of cytokines, such as IFNγ and IL-12, resulting in a Th1-mediated cellular response (42). Although the recognition of C. albicans by TLR4 has been shown in vitro and in vivo, differences between various Candida strains may account for reports showing that TLR4 plays a minor role in candidiasis (43–45). Indeed, Bellocchio et al. (46) have reported variable results, with TLR4–/– mice showing a defective Th1-protective immunity to C. albicans in models of intragastric infection and intravenous re-infection, particularly with respect to hyphae.
Thus, it is currently discussed that predominantly TLR2, but not TLR4, plays a significant role in protecting the host against primary C. albicans infection (44). Whereas Netea et al. (33), however, argue that TLR2 confers host susceptibility to infection, and while yeast cells signal through both TLR2 and TLR4, hyphae signal only through TLR2 (32).
TLR 9, an intracellular receptor, is expressed by monocytes, macrophages, DCs and CD4+ cells. Poorly methylated fungal DNA (CpG) sequences are the ligands of TLR9. Blocking TLR9 in human monocytes or utilizing TLR9-deficient mouse macrophages, stimulated with C. albicans, leads to decreased production of cytokines, mainly of IL-10 (47). Similarly, C. albicans DNA induces IL-12p40 production in bone marrow-derived myeloid DCs via TLR9 using a mechanism independent of the unmethylated CpG motif (48). Although Bellocchio et al. (46) found less production of pro-inflammatory cytokines such as IL-4, IL-10 and IL12, there was no higher mortality of TLR9–/– mice infected with C. albicans. Thus, most of the data suggest existing compensatory pathways that are mediated by other PRRs.
C-TYPE LECTIN RECEPTORS
C-type lectin receptors (CLRs) represent a family of PRRs, which are found mainly on APC. They were first defined in 1998 by the ability to bind carbohydrates in a Ca2+-dependent manner (49). These proteins are mainly membrane-bound receptors sharing one or more carbohydrate recognition domains (CRDs). Several of these CLRs can directly induce production of cytokines, while they also modulate innate and adaptive immune responses or function as binding and uptake receptors. They include several receptors implicated in fungal recognition, including mannose receptor (MR), DC-specific intercellular adhesion molecule-3-grabbing non-integrin (DC-SIGN), dectin-1 and -2, mincle, galectin-3, mannose-binding lectin, surfactant protein a (SP-A), surfactant protein D (SP-D), langerin and complement receptor 3 (CR3).
Mannose receptor
The MR is a CLR. It is expressed mainly by macrophages, as well as by DCs. Several organisms, including C. albicans, C. neoformans, Pneumocystis carinii, and other pathogens, such as bacteria and viruses, are recognized by the MR (50, 51). The receptor binds mannose, fructose, N-acetylglucosamine, and glucose (11). After recognition the receptor mediates the internalization of pathogens leading to intracellular killing. The detailed role of fungal phagocytosis has been addressed in several studies (51, 52). Yet the ability of the MR to mediate phagocytosis has been questioned (53). Because MR has no known cytoplasmic signalling motif, little is known about the signal pathways activated by engagement of MR. However, MR mediates a variety of cellular responses, including induction of NF-κB activation and the production of numerous defensive cytokines. Cross-linkage of MR with an activating anti-MR-antibody has been shown to induce cytokine (IL-10 and IL-1Ra) production in DC, and up-regulation of co-stimulatory molecules leading to Th2 polarization in vitro (54). In murine models binding of MR by C. albicans induced the pro-inflammatory cytokines IL-1β, IL-6, and GM-CSF (55), and MR deficiency led to a reduction of TNF-α and MCP-1 release in response to C. albicans uptake (56). MR recognizes branched α-linked mannose structures and N-linked mannans of C. albicans (20, 57). Some data show the effect of the MR in response to C. albicans defences. Lee and others have demonstrated that intraperitoneal injection of C. albicans induces an increased fungal burden in organs of MR–/– mice, although mortality was not influenced (58). Netea et al. (59) investigated the role of MR in mutant C. albicans strains defective in O-linked and N-linked mannans. The study demonstrated that MR recognizes N-bound mannans from C. albicans. These investigations correspond with previous studies, which have shown that MR preferentially recognizes α-linked oligomannoses (57).
Dendritic cell-specific intercellular adhesion molecule-3-grabbing non-integrin (DC-SIGN) (CD209)
DC-SIGN is specifically expressed on DCs and macrophages (60). Alpha-linked mannose structures of C. albicans are recognized by DC-SIGN. This receptor recognizes C. albicans in a calcium-dependent way and leads to uptake and phagocytosis of fungus (61). Studies have shown that DC-SIGN activates intracellular signal pathways involving Raf-kinase, ERK1 and ERK2, leading to cytokine production and modulation of TLR signal pathways (62). Activation of these signal pathways leads to an enhanced IL-10 production, thereby favouring a Th2 cellular immune response as demonstrated by Gringhuis et al. (63) and Caparros et al. (64). Stimulation of DC-SIGN signal pathways for immunomodulation in vaccination is therefore likely to elicit mainly Th2 responses, limiting the usefulness of DC-SIGN as target for adjuvant effects (65).
Dectin-1
Dectin-1 is a PRR that plays an important role in antifungal innate immunity. Dectin-1 is expressed on phagocytes (monocytes, macrophages, DCs) and neutrophils. Dectin-1 (like CR3) recognizes ß-(1,3)-glucans, which are localized on the outer layer of C. albicans yeast forms. After filamentous transformation to the hyphae variant ß-glucan remains in the inner layer of the fungus, and C. albicans can escape recognition via dectin-1 (66). Dectin-1 is a type II transmembrane protein with a C-type lectin-like CRD connected to the transmembrane region, followed by a cytoplasmic region. After binding to its ligand, dectin-1 is phosphorylated by the kinase Syk. Activated-Syk induces CARD9, which leads to IL-2 and IL-10 production in DCs and development of CD4+ IL-17-producing effector T cells (TH-17 cells) (67). Data suggest that dectin-1 induces Syk-independent signalling mediated by serine-threonine kinase RAF-1, which is suggested to activate NFκB and regulates expression of IL1ß, IL-6, IL-10 and IL-12 (68).
Several facts suggest that dectin-1 plays an important role in mucosal immunity against Candida (65). First, dectin-1 is expressed in the intestinal tract, and the outgrowth of Candida in the digestive tract from dectin-1−/− mice has been shown to be disproportionally high, leading to occlusion and contributing to the increased mortality in this study (69). Also, dectin-1 is paramount for IL-17 induction by Candida (67, 70), and patients with an impaired IL-17 production caused by STAT3 mutations (hyper-IgE syndrome) and chronic mucocutaneous candidiasis have recurrent Candida infections (71, 72).
Dectin-2
In contrast to dectin-1, dectin-2 is a receptor that is unable to induce a signalling pathway by itself. Dectin-2 is expressed on macrophages and DCs and binds C. albicans hyphae. The exact structure of the dectin-2 hyphal ligand is still not known, but has been suggested to be a mannan-like or mannan-containing glycoprotein (68). Sato et al. (73) found that the Fc receptor γ (FcRγ) chain can bind to dectin-2, which leads to phosphorylation of FcRγ. FcRγ induces activation of NFκB and the production of TNF-α and IL-1 receptor antagonist and finally leads to phagocytosis. Robinson et al. (74) reported that specific monoclonal antibodies (mAb) against dectin-2 cause decreased production of Il-2, Il-10 and TNF and are capable of suppressing the T-cell production of Candida-specific Th17 cells in cooperation with dectin-1. Similarly, another study suggests that dectin-2 is important in host defence against C. albicans by inducing Th17 cell differentiation. In particular, the yeast form of C. albicans induced Il-1ß and Il-23 secretion in a dectin-2-dependent manner (74). Bi et al. (75) have postulated that hyphae-induced activation of NFκB through CARD9/Syk-pathway is mainly due to dectin-2, but not dectin-1.
Although many receptors are involved in the recognition of the two forms of C. albicans, dectin-1 and dectin-2 are unique in providing a form-specific recognition: dectin-1 recognizes yeast through binding of β-glucan, while dectin-2 recognizes the hyphae through binding to a high-mannose-oligosaccharide that remains unidentified. Using dectin receptors, DC may distinguish yeast from hyphae, whereafter diverse signalling pathways may adjust DC function to better combat each of these forms (68).
Mincle
Mincle (also called Clec4e and Clecsf9) represents another CLR. Among the CLRs it belongs to the dectin-2 family (76). Mincle is expressed predominantly on macrophages and is not a phagocytic receptor for C. albicans. Wells et al. (76) reported that Mincle is able to mediate inflammatory responses to the yeast. Mice deficient in Mincle showed a significantly increased particular susceptibility to systemic candidiasis. In addition, mice deficient in NF-IL-6 transcription factor, which is known to control Mincle expression, are also particularly susceptible to infection with Candida (77). Different results were obtained by Graham & Brown (78) who used a different strain of C. albicans. They postulated that Mincle specifically recognizes mannose on the fungal surface. In contrast to other fungi the group could not demonstrate the recognition of C. albicans by Mincle.
Galectin-3
Galectin-3 is identical to Mac-2 (macrophage galactose-specific lectin-2). This CLR is expressed mainly by macrophages. Truong et al. (79) reported expression of galectin-3 on human eosinophils. Galectin-3 binds to ß-1,2-oligomannans isolated from C. albicans (80). Another study has demonstrated a crucial role for galectin-3 in the recognition of yeast in the gut: macrophages sense C. albicans through a mechanism involving TLR2 and galactin-3, which probably bind a ligand specific to the C. albicans cell wall surface: ß-1,2 mannosides (81). The absence of galectin-3 reduces chemically-induced inflammation in the gut of mice and abolishes the response of TLR-2 and TNF to C. albicans colonization (82).
Langerin
Langerin is a CLR that is strongly expressed by Langerhans’ cells (LCs) a subset of DCs localized in epithelium of epidermis and mucosal tissues. Langerin is a type II transmembrane protein, which recognizes pathogens and initializes several immune responses against invading pathogens. In 2002, Takahara and others showed that murine langerin is able to bind C. albicans (83, 84). Stambach & Taylor (85) reported recognition of the cell wall structures mannose, fucose and N-acetyl-glucosamine monosaccharides (GlcNAc) by langerin. De Jong (86) and others demonstrated that langerin recognizes not only carbohydrate, but also interacts with mannan and ß-glucan structures of C. albicans. Langerin’s precise immunological role, however, remains largely unknown. It has been implicated to play an important function in HIV-1 infection and defense (68).
COLLECTINS
The collectins represent a unique group of 5 human and 6 mouse proteins secreted to form soluble multimeric complexes of receptors. A total of 3 collectins, mannose-binding lectin (MBL) and surfactant proteins A (SP-A) and D (SP-D), have been shown to be involved in a wide range of the innate immune responses, including antifungal host defence (68). Mannose-binding lectin (MBL) is secreted by the liver and can bind fungus including C. albicans. MBL-deficient mice infected with C. albicans have been demonstrated to have a higher mortality than normal animals (87). SP-A and SP-D are expressed mainly in the lung and also recognize C. albicans (88).
Rosseau et al. (89) reported that SP-A down-regulated the pro-inflammatory cytokine response of alveolar macrophages and monocytes to C. albicans, including TNF, IL-1ß, macrophage inflammatory protein-1α, and monocyte chemoattractant protein-1. In another study the same group showed that SP-A has no effect on phagocytosis of C. albicans (90).
SP-D interaction with C. albicans appears to be more effective for antimycotic host defence. Van Rozendaal et al. (91) found that SP-D binds C. albicans. Moreover, the group demonstrated that SP-D inhibits phagocytosis of C. albicans by alveolar macrophages. The mechanisms by which SP-A and SP-D promote microbial agglutination and modulate phagocytosis and cytokine production, however, are unknown. Interestingly, the type of biological response elicited by the surfactant proteins appears to depend largely on the specific pathogen with which they interact.
MBL binds yeast- and pseudohyphal- growth-forms of C. albicans. Van Asbeck et al. (92) reported that C. albicans is a potent activator of the lectin pathway for complement activation. Addition of MBL to MBL-deficient serum increases the deposition of C4 and C3b and enhances the uptake of C. albicans. In another study, binding of MBL to C. albicans led to decreased phagocytosis by macrophages (93). The results suggest that both yeast cell-bound MBL and macrophage-bound MBL may be important modulators of C. albicans interactions. Ghezzi et al. (94) reported that MBL interaction with C. albicans leads to an increased production of TNF. In the mouse model low levels of serum MBL are associated with susceptibility to several diseases (94). Another study showed that double KO MBL A and C mice were more susceptible to intraperitoneal and intravenous infection with C. albicans compared with normal mice (95).
Different studies reported the role of MBL in vulvovaginal candidiasis. Increased densities of MBL were found in case of C. albicans infection. In addition, women with recurrent vulvovaginal candidiasis showed increased levels of cervicovaginal MBL (96–100).
COMPLEMENT RECEPTOR 3
Complement receptor 3 (CR3) is a widely expressed ß2-integrin consisting of CD11b and CD18. CR3 recognizes different endogenous and exogenous ligands. It is a phagocytic receptor, which is strongly expressed by neutrophils. One natural ligand of CR3 is ß-glucan, which is recognized by a lectin-domain of CR3. The lectin domain is able to bind the yeast and the hyphal growth form of C. albicans (101–104). In particular, the recognition of the minor cell wall component ß-(1,6)-glucan appears to be mediated by CR3 (105). Binding of CR3 does not mediate protective host response, but can be exploited for suppressing pro-inflammatory signals, such as TNF-release (106).
NOD-LIKE RECEPTORS
NOD-like receptors (NLRs) are a family of intracellular immune receptors characterized by leucine-rich repeats and a nucleotide-binding domain (11). In addition to the mostly cell membrane-bound TLRs and CLRs, the mammalian host defence has created a second line of recognition receptors located in the cytoplasm that, in the case of C. albicans, is represented by receptors of the NLR family (40). A significant role for anti-Candida host defence has been proposed for the NLRP3 (NLR family pyrin domain containing 3) inflammasome (40). These large multi-protein complexes activate, upon recognition of a microbial PAMP, caspase-1, leading to the processing and secretion of IL-1β and IL-18 (107). Reports link IL-1β production induced by C. albicans to the NLRP3 inflammasome (11, 108–110). Importantly, TLR2, dectin-1, and NLRP3 were all crucial for the protection from dissemination of C. albicans in a murine model of oral mucosal infection (109). However, to date, the extent to which oral epithelial cells contribute to the observed defence mechanism is not known.
CONCLUSION
During the past few years it has become increasingly clear that PRRs are essential for the host response to C. albicans, with various TLRs and LRs having distinctive roles in innate immunity. Each ligand–receptor system activates specific intracellular signalling pathways, which in turn leads to modulation of various components of the host immune response. While some receptors, such as TLR4, dectin 1 and the MR, exert a more pro-inflammatory role, others wield immunosuppressive effects (for example, TLR2, CR3 and FcγR). After discovery and defined elucidation of the role of TLRs in fungal recognition, further studies have elucidated the role of the C-type lectin receptors with a focus primarily on dectin-1 and dectin-2.
Many investigations have been conducted on this important topic, particularly with C. albicans, and thus we have gained a much better understanding of the relevant structures of the PAMPs and PRRs. However, further studies are needed in order to achieve insight into the complex interaction between PAMPs and the corresponding receptors. Indeed, co-stimulation via multiple PAMP–PRR interactions may increase both the sensitivity and the specificity of the immune recognition process. A detailed understanding could potentially provide options for novel therapeutic strategies, leading to a more targeted and successful treatment or prophylaxis of fungal infections. Also, the choice of the respective experimental model and C. albicans strain utilized appears to have significant impacts on the outcome of infection. Further research is needed to fully elucidate the role of PRRs in the various types of C. albicans infections in vivo. Many studies on disseminated candidiasis have been undertaken, but, so far, little work has been published for other forms of C. albicans infection, such as oropharyngeal or vaginal candidiasis.
ACKNOWLEDGEMENT
The authors would like to thank Monika Döbl for her assistance in manuscript preparation.
REFERENCES

