


1Lee Kong Chian School of Medicine, Nanyang Technological University, Singapore, Singapore, 2Department of Biomedical Sciences, Copenhagen Wound Healing Center, Bispebjerg Hospital, University of Copenhagen, Copenhagen, Denmark, and 3Division of Dermatology and Venereology, Department of Clinical Sciences, Lund University, Lund, Sweden
#These authors contributed equally.
Carbonic anhydrases are ubiquitously expressed enzymes that reversibly hydrate carbon dioxide to bicarbonate and protons. While the main function of carbonic anhydrases is to regulate pH and osmotic balance, their involvement in other physiological processes remains to be explored. This study analysed changes in mRNA and protein levels of carbonic anhydrase II in human primary keratinocytes treated with various toll-like receptor agonists and cytokines. A significant upregulation of carbonic anhydrase II at the mRNA and protein levels was observed upon treatment with polyinosinic-polycytidylic acid, a toll-like receptor 3 agonist. Furthermore, in agreement with the increased expression of carbonic anhydrase II in atopic dermatitis skin, carbonic anhydrase II was upregulated by the Th2 cytokines interleukins -4 and -13. In conclusion, these results suggest a potential role of carbonic anhydrase II in Th2-dependent and toll-like receptor 3-induced pathways in inflammatory skin conditions.
Key words: carbonic anhydrase II, keratinocytes, TLR3, poly(I:C), Th2 cytokines, inflammation.
Accepted May 4, 2018; Epub ahead of print May 8, 2018
Acta Derm Venereol 2018; XX: XX–XX.
Corr: Bani Kaur Suri, Lee Kong Chian School of Medicine, Nanyang Technological University, Experimental Medicine Building, 59 Nanyang Drive, Singapore 636921, Singapore. E-mail: banisuri@ntu.edu.sg
Carbonic anhydrases (CAs) reversibly convert carbon dioxide and water to bicarbonate and protons, thereby regulating pH and osmotic balance in different tissues. However, their role in skin health is unclear. Increased levels of a CA isoform, CA II, were found in human keratinocytes upon treatment with a toll-like receptor 3 (TLR3) agonist, polyinosinic-polycytidylic acid (poly(I:C)), and Th2 cytokines, IL-4 and IL-13. The TLR3 pathway is implicated in antiviral and wound responses, and Th2 cytokines are involved in eczema. This suggests that CA II has a role in inflammatory skin conditions.
Skin forms the first line of defence and thus is an important site for immune response. Toll-like receptors (TLRs) constitute a family of signal transducing proteins, which sense the environment and activate downstream pathways related to inflammation, antiviral response, maintenance of homeostasis and regeneration (1). While TLRs play a crucial role in the cutaneous innate immune system, cytokines form part of the adaptive immune response by inducing and executing immune and inflammatory responses.
Carbonic anhydrases (CAs) catalyse the inter-conversion of carbon dioxide and bicarbonates with the release of a proton (H+). Due to this fundamental, yet crucial reaction, CAs are involved in regulating pH, osmotic and electrolytic balance across kingdoms (2). Of the 15 CAs identified in humans, very few have been reported in keratinocytes and even fewer have their functions characterized. Carbonic anhydrase II (CA II), first discovered in red blood cells, has also been reported in keratinocytes. Although involvement of CA II in regulating pH in the skin has been hypothesized, its role is still unclear (3).
To investigate the regulation of CA II in the context of host pathogen interaction and the ensuing immune response, an expression study was conducted for CA II in human keratinocytes using a panel of TLR agonists and cytokines.
Adult normal human epidermal keratinocytes (NHEK) (ATCC, Manassas, USA) were cultured in dermal cell basal media, supplemented with keratinocyte growth kit, as per the ATCC recommendations. NHEK cells in plates were treated with TLR 1–9 agonists (InvivoGen, San Diego, USA) and cytokines (Milteny Biotec, Gladbach, Germany) for 12 h. Quantitative real-time PCR (qRT-PCR) was conducted for studying changes in CA2 gene expression upon treatment. Primers used were 5’-CCTGAATCCTTGGATTACTGGAC-3’ and 5’-TCCCCATTGAAGTTAAGTTTACG-3’ for CA2 and 5’-TCGACAATGGCAGCATCTAC-3’ and 5’-ATCCGTCTCCACAGACAAGG-3’ for the housekeeping gene RPLP0. Protein levels were evaluated by Western immunoblotting using CA II (ab124687, Abcam, Cambridge, UK) and RPLP0 (11290-2-AP, Proteintech, Chicago, USA) antibodies. Unbiased protein expression was quantified using an automated microscopy-based High Content Analysis (HCA) system (IN Cell Investigator 2200, GE Healthcare, New York, USA). High magnification images were captured to visualize the localization of CA II protein using confocal laser scanning microscope (LSM800, Zeiss, Jena, Germany). All experiments were repeated at least 3 times.
To investigate the regulation of CA II expression in human epidermal keratinocytes, NHEK cells were treated with a panel of TLR agonists and cytokines for 12 h. qRT-PCR analysis showed a significant up-regulation of CA2 mRNA levels following treatment with poly(I:C), a TLR3 agonist. Other tested agonists resulted in no significant changes in CA2 expression (Fig. 1A). Upon treatment with the panel of cytokines, only interleukin (IL)-4 and IL-13, both Th2 cytokines, showed a significant increase in CA2 mRNA levels (Fig. 1B). Other tested cytokines did not yield any significant changes in CA2 mRNA in primary keratinocytes.
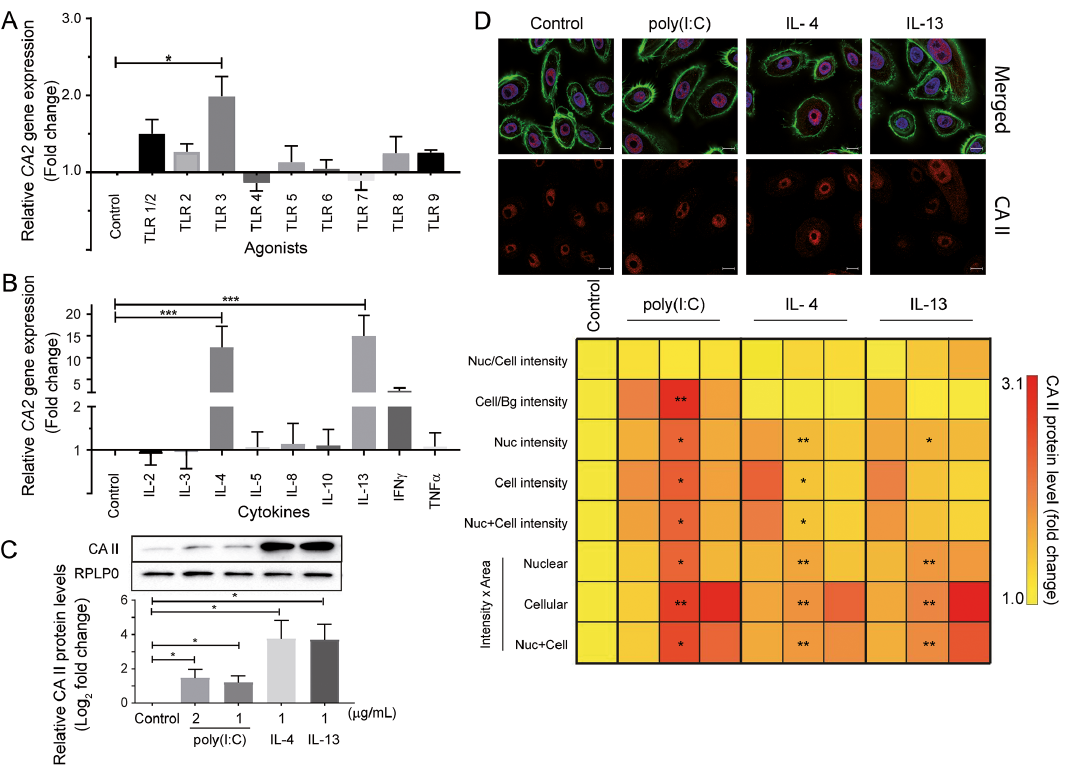
Fig. 1. Effect of toll-like receptor (TLR) agonists and cytokines on carbonic anhydrase (CA) II expression in normal human epidermal keratinocytes (NHEK) cells. NHEK cells were treated with: (A) TLR agonists; or (B) cytokines (1 μg/mL each) for 12 h in 24-well plates. Cells were rinsed and harvested for RNA extraction and subsequently subjected to cDNA synthesis. Data from quantitative real-time PCR (qRT-PCR) for CA2 mRNA levels was plotted after normalizing with RPLP0 (fold change). (C) Cells were treated with polyinosinic-polycytidylic acid (poly(I:C)) (1 μg/ml, 2 μg/ml), interleukin (IL)-4 or IL-13 (1 μg/ml) and carbonic anhydrase (CA) II protein levels were evaluated by Western immunoblotting. Relative quantitation from 3 independent experiments has been plotted with a representative immunoblot above the graph. (D) NHEK cells were treated with the TLR3 agonist poly(I:C) (2 μg/ml) and Th2 cytokines (1 μg/ml) for 12 h in 96-well plates and coverslips. Cells were rinsed, followed by immunofluorescent labelling of CA II, nucleus and cytoskeleton. High content analysis (HCA) was performed to quantify differences in protein levels and localization with respect to untreated cells. Unbiased quantitation from 27 fields of view is presented per column in a colour-coded heatmap as fold changes in CA II protein levels as captured by HCA. Representative confocal images are overlaid on the heatmap showing CA II (red), nucleus delineated with DAPI (blue) and F-actin stained with phalloidin (green). Images were captured at 63× magnification and scale bar depicts 10 µm. All data is mean ± standard deviation from 3 independent experiments. Significance was calculated by one-way analysis of variance (ANOVA) for qRT-PCR and HCA data, and by unpaired t-test for Western immunoblot data. *p < 0.05, **p < 0.01, ***p < 0.001 compared with untreated control. Nuc: nuclear; Cell: cellular: Bg: background.
The TLR3-mediated increase in CA2 gene expression was translated to a similar increase in CA II protein levels (Fig. 1C). Western immunoblot analysis showed significantly increased levels of CA II protein in human keratinocytes upon treatment with poly(I:C) and the Th2 cytokines, IL-4 and IL-13. Unbiased and large scale cell-by-cell quantitation of CA II protein in NHEK cells was carried out by HCA, which also showed a significant increase in CA II protein expression in the cytoplasm and nucleus. Visualization of the elevated CA II levels can be observed in the representative confocal images (Fig. 1D).
This study demonstrates that poly(I:C), a commonly used TLR3 agonist, induces CA II expression in keratinocytes. Keratinocytes have been reported to express TLR1, TLR2, TLR3 and TLR5; however, there have been mixed reports about the presence of TLR4, TLR6 and TLR9 (4, 5). To circumvent the discrepancies, we tested the effect of a panel of TLR agonists on cultured human epidermal keratinocytes. Amongst the TLR agonists tested, only TLR3 lead to an up-regulation of CA II expression at both mRNA and protein levels. TLR3 is located mostly in the endosomes and requires the agonist to enter the cell to initiate a cellular response. Typically, TLR3 senses virus-derived patterns triggering antiviral genes, such as type I interferons, by a myeloid differentiation factor 88 (MyD88)-independent pathway, which leads to nuclear factor (NF)-κB or interferon regulatory factor (IRF)-3 dominated immune response (6, 7).
TLR3 has also been implicated in sensing tissue damage and injury, and activating downstream repair mechanisms. Injury to skin by ultraviolet (UV) radiation and other factors releases double-stranded (ds) RNA, which activates TLR3 (8). This activation triggers epidermal barrier repair by increasing expression of corneodesmosins, junction proteins and cross-linking enzymes, such as transglutaminases and sphingomyelinases (9). Topical application of poly(I:C) has been found to augment wound closure of laser-induced wounds (10). Activation of TLR3 maintains keratinocytes in a less differentiated state contributing to skin and hair follicle regeneration (11). Induction of psoriasin (S100A7) via TLR3, which in turns promotes wound closure, has also been reported (12). Furthermore, decreased expression of TLR3 in adults compared with infants alludes to the reduced ability for repair and regeneration with ageing (13). It is notable that poly(I:C) is a synthetic dsRNA analogue, which stimulates anti-viral response elements or dsRNA sensing molecules present in keratinocytes, such as protein kinase R (PKR), RNA helicases, retinoic acid-inducible gene I (RIG-I) and melanoma differentiation associated protein 5 (MDA5), while poly(I:C)-induced IRF-3 activation and IFN-β expression do not decrease upon inhibition of TLR3 signalling (14). In this context, it is possible that the observed effect of poly(I:C) could be a combined effect of PKR, RIG-I, MDA5 and TLR3. Interestingly, foscarnet and uracils, antiviral drugs and their derivatives, inhibit the activity of a variety of CAs in vitro (15, 16) and CA inhibitors enhance the activity of a nucleo-polyhedrovirus (17). Gel-based recombinant CA IV application to full-thickness skin wounds in mice has been found to significantly improve wound re-epithelialization (18). Linking these data reported earlier with our current observations suggests that CA II is downstream of poly(I:C)-induced pathway affecting both antiviral response and wound healing.
Th2 dominant atopic dermatitis (AD) lesions show a significant increase in CA2 gene expression compared with Th1-dominant psoriatic lesions and controls (19). Our data also shows an up-regulation of CA II upon treat-ment with IL-4 and IL-13, both Th2 cytokines, which is in accordance with previously reported results (3). Absence of a genetic association of the CA2 gene with AD (20) suggests that upregulation of CA II is an effect rather than a cause of AD. Interestingly, thymic stromal lymphopoietin (TSLP), which is also upregulated in AD lesions and promotes a Th2 response (21), is also inducible by poly(I:C) (22).
One limitation of the current study is that the observations were recorded 12 h post-treatment. Upregulation of CA II mRNA and protein could be an indirect effect of poly(I:C)-induced secreted cytokines, such as IL-6, TNF-α (23), CXCL8, CXCL9, CXCL10, CCL2, CCL20, and CCL27 (24). While TNF-α and IL-8 (CXCL8) did not directly affect CA2 expression in NHEK cells (Fig. 1B), possible effects of other cytokines and/or signalling mediators that could be induced in human keratinocytes via TLR3 cannot be ruled out. It is also possible that other dsRNA sensing receptors are operative in poly(I:C)-induced upregulation of CA II in human keratinocytes. Another point to note is that this study used a relatively high concentration of cytokines (1 μg/ml) to confirm the expression of CA II upregulation in human keratinocytes. Nevertheless, we present novel data of CA II upregulation in poly(I:C)-treated human keratinocytes when stimulated directly or indirectly via TLR3. It would be interesting to comprehensively screen the potential involvement of relevant cytokines/chemokines and their combinations through time-course and dose-course studies using a physiological concentration range.
In conclusion, this study shows that the TLR3 agonist poly(I:C) upregulates CA II in human keratinocytes, which could have implications for antiviral, wound healing and/or regeneration pathways. Further research is needed to comprehensively characterize CA II for its potential role in inflammatory and skin proliferative disorders.
The authors acknowledge start-up grants to NKV and AS provided by Lee Kong Chian School of Medicine, Nanyang Technological University, Singapore and Singapore Ministry of Education, Academic Research Fund Tier 1 (Project No. 2015-T1-001-082) for funding this research.
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize