


1Dermatology Unit, Department of Medicine Solna, Karolinska Institutet, 2Center for Molecular Medicine, Department of Molecular Medicine and Surgery, Karolinska Institutet, SE-171 76, Stockholm, Sweden, and 3Department of Dermatovenereology, Faculty of Medicine, Gondar University, Gondar, Ethiopia. E-mail: samina.asad@ki.se
Accepted Sep 23, 2018; Epub ahead of print Sep 24, 2018
Atopic dermatitis (AD) is an inflammatory skin disorder characterized by pruritus and dryness of the skin in typical locations. The incidence rate of AD varies worldwide. In Europe the prevalence is up to 20% in children, while reports in Ethiopia vary between 1% and 2% in rural areas and 19% in urban areas (1, 2).
The major risk gene for AD in Europeans is the filaggrin (FLG) gene (1q21), where a number of null-mutations as well as variations in gene dose are associated with disease susceptibility (2). FLG mutations have also been identified in the Asian population, although these mutations seem to be more population- or family-specific (2). Little is known about AD susceptibility genes in the African population. Our previous studies indicate a low prevalence of FLG null-mutations in Ethiopian cases of AD (1). This is in line with studies performed in the South African AmaXhosa population, suggesting that, in the African population, genes other than FLG may be involved in the development of AD (3).
In searching for risk genes for AD in the Ethiopian population, we previously performed whole-exome sequencing (WES) of 22 Ethiopian patients with AD/ichthy-osis vulgaris in whom several genetic polymorph-isms were detected (1).
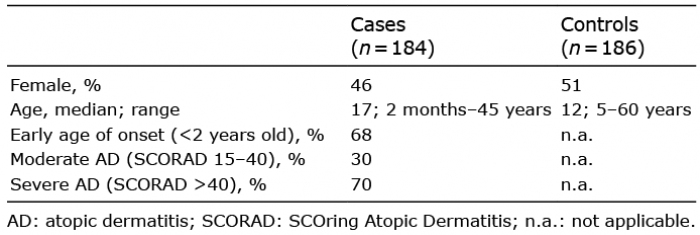
This study aimed to elucidate whether some of the single-nucleotide variants (SNVs) identified by Taylan et al. are associated with AD susceptibility in our Ethiopian AD cohort, consisting of 184 cases of AD and 186 healthy controls (Table I) including the 22 previously sequenced individuals (1).
Table I. Ethiopian cases and controls
All study participants gave written or oral informed consent, which complies with the principles of the Declaration of Helsinki. This study was approved by the Ethics Review Board of the University of Gondar, and the regional ethics committee, Karolinska Institutet, Sweden.
All patients were diagnosed with AD according to the UK Working Party’s diagnostic criteria (2) and answered questionnaires regarding other atopic manifestations (1). The samples were obtained from ALERT Dermatology Hospital, Tikur Anbessa Hospital and Gondar University Hospital. We chose to genotype gene variants in: (i) epidermal differentiation complex (EDC) genes; cornulin (CRNN) (4) and hornerin (HRNR) (5); (ii) non-EDC genes that had either previously been associated with AD or candidate genes with a known function in the skin; (iii) gene variants that more than one individual was carrying in the WES. Based on this strategy we genotyped 38 SNVs using TaqMan (Applied Biosystems Inc., Foster City, CA, USA) in the following 10 genes; transglutaminase (TGM) 1, 3 and 5 (6), desmoglein (DSG) 1 and 3–4 (7), a variant in C11orf30 encoding for the EMSY protein (8), SPINK5 (11), hornerin (HRNR) (5) and cornulin (CRNN) (4). Primers/probes were obtained from Thermo-Fisher Scientific Inc., (Foster City, CA, USA). Genotyping indicated that 28 SNVs were non-polymorphic or did not detect minor allele homozygotes in our material (Table SI). For the remaining 10 SNVs (Table II and Table SII), association with AD was calculated using χ2 tests on both genotype and allele frequencies. Fisher’s exact test was performed in instances where the counts were lower than 5. R program v.2.12.2 was used to calculate logistic-regression using generalized linear modelling between patients and controls. SNV frequencies were checked to follow Hardy-Weinberg equilibrium.
Table II. Minor allele frequencies (MAF) in our cases-control material and for the African population in Exac-and GnomAD databases
The genotyping success and concordance rates for SNVs showing 3 genotypes was above 90%. Furthermore, all successfully genotyped SNVs (showing all 3 genotypes) correlated with the previous whole exome-sequence results.
In our WES study, 12 SNVs were identified in the HRNR gene, which is a component of the epidermal cornified cell envelope thought to share similar features with filaggrin and demonstrated to have reduced expression in the epidermis of patients with AD (5). In the HRNR gene, only 3 SNVs demonstrated 3 genotypes and were analysed further. Two SNVs were analysed in CRNN, which is a marker of late epidermal differentiation and down-regulated in AD skin (4). However, these SNVs were non-polymorphic in our study material and discarded from further studies. In addition, SNVs were genotyped in DSG1-4 and TGM1, 3 and 5, all of which have important roles in forming the epidermis and are associated with altered expression in AD skin (6, 7). One SNV located in the c30orf11 region, which has been significantly associated with AD in previous studies, was also analysed (8). In the SPINK5 gene, 7 SNVs were genotyped. SPINK5 (5q32) encodes for the skin barrier protein Lympho-epithelial Kazal-type-related inhibitor (LEKTI), which plays a major role in controlling the proteolytic processing of profilaggrin into FLG monomers. Loss-of-function mutations of SPINK5 cause the rare autosomal recessive disorder Netherton syndrome characterized by congenital ichthyosis as well as manifestations overlapping with AD. SNVs in this gene have previously shown modest association with AD in European and Asian populations (9). When association was calculated only between patients with severe AD and controls, significant association was observed for both rs230363 (p = 0.023) and rs2303067 (p = 0.025) in SPINK5. These findings are in line with previous studies in European and Asian populations (9). This prompted us to perform immunohistochemistry with the SPINK5 antibody (Novus Biologicals, Abingdon Oxon, UK), on skin biopsies of Ethiopian patients with AD. Similar staining signals were indicated in a rs230363 homozygous wild-type carrier a homozygous minor allele-carrier and a heterozygous carrier (Fig. S1). However, we did not find any association when logistic regression was calculated. Association was also tested in all SNVs for age at onset (ages < 2, 5 and 8 years, respectively) vs. controls and for AD severity measured by SCOring Atopic Dermatitis (SCORAD).
In our study, most SNVs had low minor allele frequencies (MAF) corresponding to frequencies in the African population in the online databases ExAc and GnomAD (Table II and Table SI) and, therefore, since our cohort is small, we may be limited to identify all 3 genotypes for many of the SNVs. Although it may be concluded that, due to low MAF, none of the variants would have a large effect on risk of AD. On the other hand, since most of the genes analysed are known to have altered expression in patients with AD compared with healthy controls, a dose-dependent or change of expression of the genes in the Ethiopian population that lead to AD cannot be ruled out.
Bearing in mind that FLG is not associated with AD in the Ethiopian population and no other major disease-susceptibility genes have been identified so far these results indicate that more genetic studies need to be carried out in order to understand the genetic background of AD in the Ethiopian population.
All patients and healthy control individuals attending the Department of Dermatovenereology, Faculty of Medicine, Gondar University, Gondar, Ethiopia. Supported by Edward Welander Foundation (2206) and the Swedish Asthma and Allergy. Association’s Research Foundation (2013040) and through the regional agreement on medical training and clinical research (ALF) between Stockholm County Council and Karolinska Institutet.
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize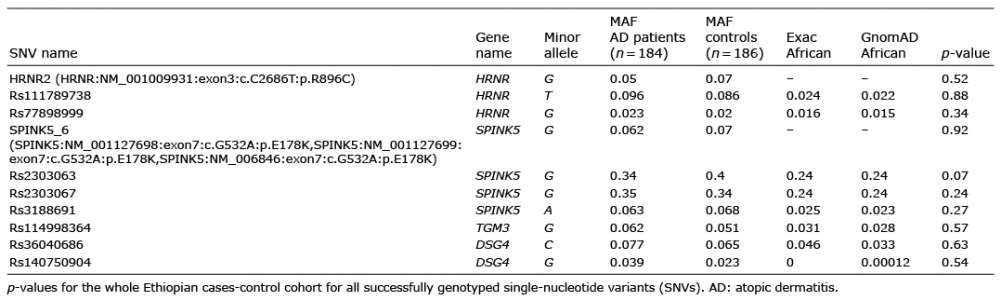 Click to show fullsize
Click to show fullsize