


1Data Analysis Unit and 2Dermatology Department, La Princesa University Hospital, La Princesa Health Research Institute (IIS-IP), 3Clinical Pharmacology Department, La Princesa University Hospital, Teófilo Hernando Institute, Madrid Autonoma University (UAM), La Princesa Health Research Institute (IIS-IP), Madrid, 4Dermatology Department, La Fe University and Polytechnic Hospital, 5Dermatology Department, La Fe University and Polytechnic, Hospital, La Fe Health Research Institute Valencia University, Valencia, 6Center of Biomedical Research Network in Liver and Digestive Diseases (CIBERehd), Carlos III Health Institute, and 7School of Pharmacy, CEU San Pablo University, Madrid, Spain
Biological drugs targeting tumour necrosis factor are effective for psoriasis. However, 30–50% of patients do not respond to these drugs and may even develop paradoxical psoriasiform reactions. This study searched for DNA copy number variations that could predict anti-tumour necrotic factor drug response or the appearance of anti-tumour necrotic factor induced psoriasiform reactions. Peripheral blood samples were collected from 70 patients with anti-tumour necrotic factor drug-treated moderate-to-severe plaque psoriasis. Samples were analysed with an Illumina 450K methylation microarray. Copy number variations were obtained from raw methylation data using conumee and Chip Analysis Methylation Pipeline (ChAMP) R packages. One copy number variation was found, harbouring one gene (CPM) that was significantly associated with adalimumab response (Bonferroni-adjusted p-value < 0.05). Moreover, one copy number variation was identified harbouring 3 genes (ARNT2, LOC101929586 and MIR5572) related to the development of paradoxical psoriasiform reactions. In conclusion, this study has identified DNA copy number variations that could be good candidate markers to predict response to adalimumab and the development of anti-tumour necrotic factor paradoxical psoriasiform reactions.
Key words: pharmacogenomic; CNV; psoriasis; anti-TNF drug; methylation.
Accepted Apr 1, 2021; Epub ahead of print Apr 13, 2021
Acta Derm Venereol 2021; 101: adv00448.
doi: 10.2340/00015555-3794
Corr: Esteban Daudén, Dermatology Department, Francisco Abad-Santos and María C. Ovejero-Benito, Clinical Pharmacology Department, La Princesa University Hospital, Teófilo Hernando Institute, Madrid Autonoma University (UAM), La Princesa Health Research Institute (IIS-IP), Diego de León 62, ES-28006 Madrid, Spain. E-mails: estebandauden@gmail.com; francisco.abad@salud.madrid.org; maria.ovejerobenito@ceu.es
Currently, 30–50% of patients with psoriasis do not respond to anti-tumour necrotic factor drugs, moreover, some patients develop complications. The aim of this study was to determine biomarkers that can predict drug response and complications, such as psoriasiform reactions. Bioinformatic tools were used to compare sections of the genome that are repeated (copy number variations), which could contain genes. Copy number variations were identified that could predict response to adalimumab. Moreover, copy number variations were found that could predict the development of psoriasiform reactions. These results may enable therapies to be improved. The biomarkers may help clinicians to avoid complications derived from anti-tumour necrotic factor drugs, and to optimize patients’ care.
Psoriasis is a chronic inflammatory disease with an important genetic component (1). In addition to affecting the skin, psoriasis is associated with other pathologies, such as psoriatic arthritis, increased cardiovascular risk, Crohn’s disease, and depression (2)
Autosomal genes present 2 copies, one from each of the progenitors. Copy number variations (CNVs) represent a decrease (deletion) or increase (duplications or insertion) in the number of copies of a DNA region (3). CNVs can affect gene expression by altering the genome structure and transposable elements within a region. Although CNVs are present in healthy individuals, they can be involved in different pathologies, such as cancer and autoimmune diseases (4). CNV determination is currently being applied to the diagnosis of various diseases (5).
The role of CNVs in psoriasis has been shown in several studies (6–8). An association has been described between psoriasis and the homozygous or heterozygous deletion of the LCE3B and LCE3C genes, which are part of the cluster of late cornified envelope genes (LCE) involved in the terminal differentiation of the epidermis (7). Moreover, an increase in copy number of the CCL4/CCL4L genes, which encode for chemokines that stimulate T helper type 1 (Th1) cells, T regulatory (Treg) cells, monocytes and dendritic cells (DC), is related to greater severity of psoriasis (6). Psoriasis is also linked to CNVs in β-defensin (DEFB) genes, which encode cationic antimicrobial peptides with an important role in both innate and adaptive immunity (8).
Anti-TNF drugs are effective in treatment of moderate-to-severe plaque psoriasis. However, 30–50% of patients do not present an adequate response (9). Moreover, these drugs are expensive and, although rarely, patients can develop adverse effects, including paradoxical psoriasiform reactions (10, 11). Pharmacogenetics has been widely applied to the search for genetic biomarkers of drug response in psoriasis (12). Nevertheless, to our knowledge, CNV analysis has not yet been applied to pharmacogenetics in psoriasis. Thus, the aim of this study was to identify CNVs throughout the whole genome that can predict anti-TNF drug response in patients with psoriasis or the appearance of anti-TNF-induced paradoxical psoriasiform reactions.
Study population and ethics statement
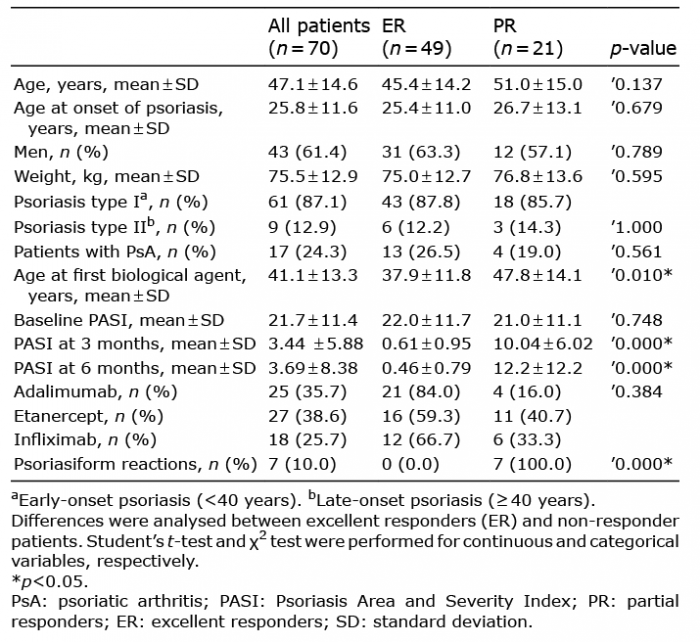
This study was designed as a non-interventional, cohort, cross-sectional observational study. Blood samples were obtained from adult patients with moderate-to-severe plaque psoriasis (according to the consensus document of the Spanish Academy of Dermatology and Venereology Psoriasis Working Group) (13), treated with anti-TNF drugs (adalimumab, infliximab or etanercept) who signed the informed consent. The protocol and informed consent document complied with Spanish legislation on biomedical and clinical research and were approved by the Ethics Committee for Clinical Research of Hospital Universitario de la Princesa. Patients exhibiting extreme phenotypic response to anti-TNF drugs were selected in order to maximize differences in drug response. Patients were distributed into 2 groups: excellent responders (ER) if they achieved PASI90 response (90% improvement from baseline Psoriasis Area and Severity Index (PASI) score) at 3 and 6 months, and partial responders (PR) if they did not achieve PASI75 response (75% improvement from baseline PASI score) (Table I, Fig. 1).
Table I. Summary of the clinical and demographic characteristics of the study population
Fig. 1. Workflow of the analysis performed in this study. CNV: copy number variations; CpG: cytosine-phosphate-guanine methylation sites; ER: excellent responders; IDAT: raw methylation files; PR: partial responders; TNF: tumour necrosis factor.
Genotyping
DNA was extracted from peripheral blood samples using the MagNa Pure® System (Roche Applied Science, Penzberg, Germany). DNA integrity number was obtained with 2100 BioAnalyzer (Agilent technologie, Santa Clara, CA, USA). The EZ DNA Methylation Kit (Zymo Research, Irvine, CA, USA) was then used for bisulphite conversion of 1,000 ng genomic DNA. Genome-wide DNA methylation analysis was performed with high-density array Illumina Infinium Human Methylation 450 Bead Chip (Illumina Inc., San Diego, CA, USA) in blood samples, according to the manufacturer’s protocol, as described previously (14). The data presented in this publication have been deposited in NCBI’s Gene Expression Omnibus (https://www.ncbi.nlm.nih.gov/geo/) and are accessible through GEO Series accession number GSE151278 (14).
Copy number variation analysis
All analyses were performed on the raw data (IDAT files) obtained from microarrays. IDAT files were analysed with 2 R packages specific for CNV analysis based on methylation microarrays: conumee and the Chip Analysis Methylation Pipeline (ChAMP) (15).
Chip Analysis Methylation Pipeline
CNVs were obtained with ChAMP using the “myCNA” function (15). ChAMP combines the intensity values of the methylated and unmethylated probes of each of the cytosine-phosphate-guanine (CpG) sites. It then normalizes the intensity values with a series of controls (genomes from healthy individuals) obtained from the minfi.Data package (minfiData: Example data for the Illumina Methylation 450k array. R package version 0.36.0). This process returns segments with a length ranging from 1,000 to over 2,000,000 base pairs (Fig. 1).
Conumee
IDAT data were pre-processed with the minfi package, followed by processing with the R package conumee (https://bioconductor.org/packages/release/bioc/html/conumee.html). This package combines the intensity values of the methylated and unmethylated probes of each of the CpG sites and normalizes the intensity values with a series of controls obtained from the minfi.Data of minfi.Data.EPIC packages. Conumee executes 2 different forms of DNA fragmentation: bins and segments. Bins are DNA regions that contain 15 neighbouring CpGs and their number is constant for each patient (15,820). Segments are clusters that present the same state of variation in the number of copies, obtained with the circular binary segmentation (CBS) algorithm. Segments are larger than the bins, their size ranges from 100,000 to 6,000,000 base pairs, and their number varies among patients (Fig. 1).
Enrichment analysis
Significant CNV regions (either bins or segments) were annotated using the bedr R library for detecting genes located on these regions. In order to identify the relationships and signalling pathways in which these genes are involved, they were subjected to enrichment analysis using Enrichr (https://amp.pharm.mssm.edu/Enrichr/).
Statistical analysis
Specific scripts were developed for comparing bins and segments obtained by different packages in order to find representative CNVs of the different comparisons performed (Fig. 1, Table I). Bins: Student’s t-test was used to compare intensity values of each bin between the different groups analysed (i.e. ER vs PR). In order to avoid biases due to differences in segment lengths the study compared only those segments that presented the same location, same length and the same initiation and end base pairs in at least 2 patients of each group (ER and PR).
Subsequenly, Bonferroni correction was applied to avoid spurious results due to multiple comparisons. Those Bonferroni adjusted p-values lower than 0.05 or marginally significant (p < 0.065) were considered.
Copy number variations associated with anti-tumour necrosis factor drug response
Initially, the study searched for CNVs that could predict anti-TNF drug response. For that purpose, a methylation analysis of 70 patients with moderate-to-severe psoriasis treated with anti-TNF drugs was performed. Of these, 49 were ER and 21 were PR (Table I). Twenty-five of the patients (35.7%) were treated with adalimumab, 27 (38.6%) with etanercept, and 18 (25.7%) with infliximab. Of the patients treated with adalimumab, 21 (84%) were ER and 4 (16%) were PR. For etanercept, 16 (59.3%) patients were ER and 11 (40.7%) were PR. Finally, 12 (66.7%) patients treated with infliximab were ER, while 6 (33.3%) were PR.
Potential differences were observed in the CNV patterns between the ER and PR groups for global anti-TNF drug response (Fig. S1). However, a detailed analysis could not find any significant difference between ER and PR to anti-TNF drugs with any of the strategies tested (ChAMP segments, conumee segments and bins) (Table II).
Table II. Analysis of copy number variations and genes associated with anti- tumour necrosis factor (anti-TNF) drug response or to the appearance of psoriasiform reactions
Subsequently, a search was performed for CNVs that could specifically predict the independent response to each anti-TNF drug (adalimumab, etanercept or infliximab). No CNVs were found that could significantly predict etanercept or infliximab response. However, 1 bin was found showing differences between ER (n = 21, Fig. S2A) and PR (n = 4, Fig. S2B) to adalimumab. This bin included one gene (Table II and Table III, Fig. S3A). The search also found 2 additional bins, harbouring a total of 4 genes, that showed a trend toward association with adalimumab response (Table II). Nevertheless, these genes were not associated with any known signalling pathway by enrichment analysis.
Table III. Summary of significanta or marginally significantb copy number variations associated with adalimumab response or the development of paradoxical psoriasiform reactions
Copy number variations associated with development of paradoxical psoriasiform reactions
In addition, a search was carried out for CNVs that could predict which patients would develop anti-TNF-induced paradoxical psoriasiform reactions. Seven patients developed these reactions. Of these, 3 patients had been treated with infliximab and 4 with etanercept. Most of the patients (n = 6; 85.6%) experienced a change in psoriasis morphology from plaque psoriasis to guttate psoriasis (a clinical form that the patients had not shown previously). One patient (14.3%) with plaque psoriasis developed an erythrodermic flare after initiating etanercept therapy.
An attempt was made to identify biomarkers for all types of psoriasiform reactions. Initially, blood samples from psoriasis patients with (n = 7) (Fig. S4B) and without paradoxical psoriasiform reactions (n = 63) were analysed (Fig. S4A) after anti-TNF drug treatment. Two 2 bins were found, harbouring a total of 4 genes, which could potentially identify which patients would develop a psoriasiform reaction (Tables II and III, Fig. S3B). Furthermore, an additional bin was found, including 2 genes, which showed a trend towards association with psoriasiform reactions (Table II).
This study used a pharmacogenomics approach to search throughout the whole genome for CNVs that could predict anti-TNF drug response in psoriasis. Multiple pharmacogenetic studies searching for predictive markers of drug response in psoriasis have focused on single nucleotide polymorphisms (SNPs) (12). Moreover, the only pharmacogenomics study following a hypothesis-free approach (16) also focused on the analysis of SNPs associated with anti-TNF drug response. Although CNVs of pharmacogenes have already been associated with patients’ response to different drugs (17), no systematic pharmacogenomics study addressing this association has been published. Therefore, to our knowledge, this is the first study evaluating the association between CNVs throughout the whole genome and the response to biological drugs in patients with psoriasis.
Globally, no significant differences in CNVs were found between ER and PR to anti-TNF drugs. Nevertheless, one bin showing differences between ER and PR to adalimumab was found (Table III). This CNV region contains one gene: CPM (Carboxypeptidase M), which is involved in the maturation of macrophages, immune cells that play an important role in psoriasis pathogenesis (18). Moreover, 2 additional bins were found that showed a trend toward association with adalimumab response (Table III). Each bin harbours 2 genes: EP400 (E1A Binding Protein P400) and SNORA49 (adjusted p-value = 0.053) and PMEPA1 (Prostate Transmembrane Protein, Androgen Induced 1) and NKILA (NF-? B interacting long noncoding RNA) (adjusted p-value = 0.063).
EP400 is a component of the NuA4 histone acetyltransferase complex involved in transcriptional activation of specific genes, mainly by acetylation of nucleosomal histones H4 and H2A, and contributes to chromatin remodelling. EP400 is associated with SMAD2-51, which is downregulated in psoriatic skin compared with healthy skin from the same patients (19).
Surprisingly, SNORA49, PMEPA1 and NKILA have not previously been associated with psoriasis or with biological drug response. Although PMEPA1 function is not well understood, this gene is believed to encode a trans-membrane protein that can bind SMAD proteins through its cytoplasmic domain and induces Akt activation (20). NKILA modulates functions involving NF-?B pathway (20). Since PMEPA1 and NKILA are located close to each other, it has been hypothesized that NKILA downregulates PMEPA1 levels (20).
None of these CNVs involve the same genes as the SNPs capable of predicting adalimumab and infliximab drug response (rs6661932 (IVL), rs2546890 (IL-12B), rs2145623 (NFKBIA), rs9304742 (ZNF816A) and rs645544 (SLC9A8) and rs1061624 (TNFR1B) (21). This reinforces the idea that SNPs and CNVs are independent complementary genetic mechanisms contributing to variability in drug response (22), thereby suggesting that the present findings describe novel biomarker candidates. No significant differences in CNVs were observed between ER and PR to etanercept or to infliximab. Accordingly, in a previous study, we found differences in methylation characteristics between ER and PR to adalimumab, but we did not find specific biomarkers of etanercept or infliximab response (14).
In addition, we searched for pharmacogenomic biomarkers of the appearance of anti-TNF-induced psoriasiform reactions. There are different types of paradoxical psoriasiform reactions: new-onset psoriasis in patients with a different pathology, worsening of a pre-existing psoriasis, and changes in the phenotype of psoriasis (10, 23). Most of the paradoxical psoriasiform reactions included in the current study consisted in a change in psoriasis morphology from plaque psoriasis to guttate psoriasis. This study characterized 1 CNV that could potentially predict the appearance of paradoxical psoriasiform reactions (Table III). This bin harbours 3 genes: ARNT2, LOC101929586, and MIR5572. ARNT2 (Aryl Hydrocarbon Receptor Nuclear Translocator 2) is involved in hypoxia response in epidermal keratinocytes (24). Both hypoxia and angiogenesis are deeply involved in psoriasis development (25). LOC101929586 is an lncRNA gene, while MIR5572 encodes a microRNA. Both types of RNA are involved in post-transcriptional regulation of gene expression.
In addition, this study identified another bin showing a trend towards association with psoriasiform reactions (adjusted p-value = 0.061). This bin harbours 2 genes: ACCS and ACCSL. ACCS (1-Aminocyclopropane-1-Carboxylate Synthase Homolog [Inactive]) encodes an enzyme that catalyses the L-vinylglycine deamination, while ACCSL is an ACCS paralog generated by a duplication event within the genome that has developed pyridoxal phosphate binding function.
A previous study in our laboratory revealed 5 SNPs (rs11209026 in IL23R, rs10782001 in FBXL19, rs3087243 in CTLA4, rs651630 in SLC12A8 and rs1800453 in TAP1) associated with paradoxical reactions. The genes presenting these SNPs are not included in any of the CNVs described in the current study. These results reinforce the notion that CNVs and SNPs are complementary, but different, mechanisms related to the development of psoriasiform reactions.
Susceptibility to psoriasis or disease severity has been associated with CNVs in certain genes such as CCL4/CCL4L (6), LCE3B and LCE3C (7) and DEFB (8). However, we have not found CNVs associated with anti-TNF drug response or with the appearance of psoriasiform reactions in the regions containing these genes. These results suggest that, although CNVs in these regions may be involved in psoriasis pathogenesis, they might not contribute to anti-TNF drug response or to paradoxical psoriasiform reactions.
Herein, we propose a strategy for the statistical analysis of CNVs with data from methylation arrays using bioinformatics tools (conumee and ChAMP) (15). Previous publications have shown that they provide analogous results to the gold-standard technique for CNV analysis (CGH arrays) (26, 27). Nevertheless, these tools focus on CNV representation (15, 26, 28) and are not designed for the statistical analysis of differences between different groups. Thus, we have developed a strategy that could help to maximize the information gathered by expensive techniques such as methylation microarrays. Notably, this study obtained pharmacogenomic information from an array initially designed for pharmacoepigenetics analysis (14).
In order to maximize the CNV information obtained from methylation arrays this study used different R packages (ChAMP (15) and conumee). In addition, the study attempted to use other packages specific to CNV analysis of methylation data; however, these are still in development (cnAnalysis 450K (29), EpiCopy (30)) or deprecated (CopyNumber450K (31)). Meffil (32) and RnBeads (33) were discarded because they provide individual CNV information for each CpG, but do not integrate these differences to provide CNV regions. No significant differences were found in segments (with conumee or ChAMP) in any of the analyses performed. However, CNVs affecting such long DNA regions (100,000 and 6,000,000 base pairs) could cause more severe pathologies, such as some types of cancer, e.g. glioblastoma (28). Moreover, a very restrictive analysis was performed in the current study, which considered only those segments that had the same location in the genome (chromosome, start and end) and were present in most of the patients in the compared groups (i.e. ER vs PR). However, these results should be validated with other techniques in order to determine if they are dependent on method.
Study limitations
The main limitation of this study is the reduced number of patients in the different groups analysed. However, a strength of the study is that it was based on a methylation array and the sample size is higher than that used in other methylation studies (n = 12 patients) (34), (n = 24 patients) (35). Another limitation is that observed CNVs showed a small difference in the intensity values between the compared groups. The relationship between intensity values and CNV copies is unclear. However, certain publications consider a CNV gain when log2 ratio is higher than 0.2 and a loss if it is lower than –0.2 (36). Most of the reported CNVs in this study do not reach these thresholds (Table III). This could be due to heterogeneity in the peripheral blood composition, which could mask possible differences between the groups. Moreover, as this is the first study using this technique, it is necessary to validate these results using an independent cohort of patients and a different technique.
Conclusion
Through CNV analysis of methylation data, the current study has unveiled CNV regions associated with adalimumab response or psoriasiform reactions. These results could provide new insights into potential new biomarkers for these outcomes. As this is the first study to use this approach, further validation studies are required to confirm the clinical potential of these biomarker candidates.
The authors thank Guillermo Pita and Manuel Gómez Gutierrez for their valuable comments on this manuscript.
This study was supported by Instituto de Salud Carlos III PI 13/01598 and the Ministry of Science and Innovation and the European Regional Development’s funds (FEDER).
Conflicts of interest. FA-S has been a consultant or investigator in clinical trials sponsored by the following pharmaceutical companies: Abbott, Alter, Chemo, Farmalíder, Ferrer, GlaxoSmithKline, Gilead, Janssen-Cilag, Kern, Normon, Novartis, Servier, Teva, and Zambon. ED has potential conflicts of interest (advisory board member, consultant, grants, research support, participation in clinical trials, honoraria for speaking, and research support) with the following pharmaceutical companies: AbbVie (Abbott), Amgen, Janssen-Cilag, Leo Pharma, Novartis, Pfizer, MSD, Lilly and Celgene. ML-V has potential conflicts of interest as she has participated in clinical trials or as consultant with Abbvie (Abbott), Galderma, Janssen-Cilag, Leo Pharma, Pfizer, Novarties, Lilly, Almirall and Celgene. MCO-B has potential conflicts of interest (honoraria for speaking and research support) with Janssen-Cilag and Leo Pharma. The other authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript apart from those disclosed. AS-T has served as a consultant and/or paid speaker for and/or participated in clinical trials sponsored by companies that manufacture drugs used for the treatment of psoriasis, including AbbVie, Celgene, Janssen-Cilag, LEO Pharma, Lilly, Novartis and Pfizer. RB-E has served as a consultant and/or paid speaker for and/or participated in clinical trials sponsored by companies that manufacture drugs used for the treatment of psoriasis, including AbbVie, Celgene, Janssen-Cilag, LEO Pharma, Lilly, Novartis and Pfizer.


 Click to show fullsize
Click to show fullsize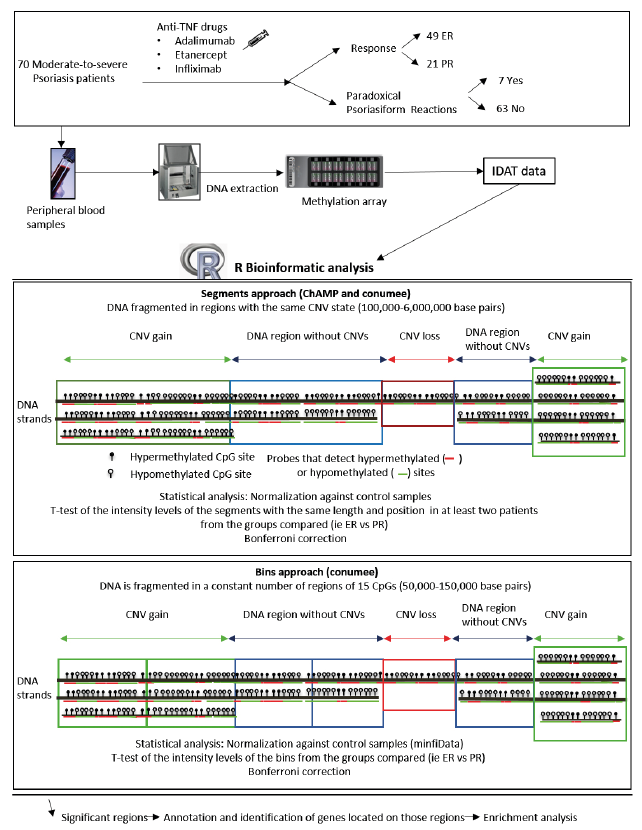 Click to show fullsize
Click to show fullsize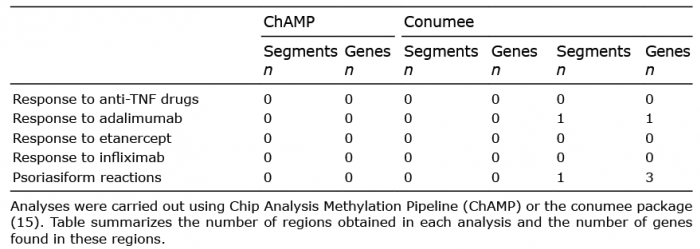 Click to show fullsize
Click to show fullsize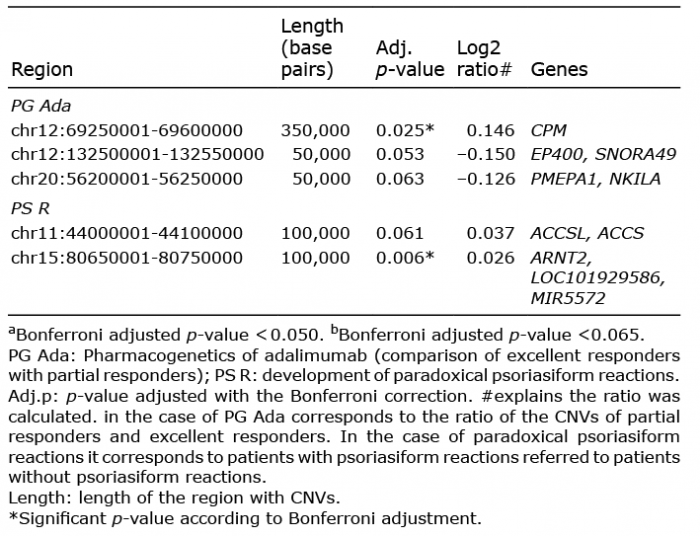 Click to show fullsize
Click to show fullsize