


1Institute for Environmental and Gender-Specific Medicine, Juntendo University Graduate School of Medicine, and 2Department of Dermatology, Juntendo University Urayasu Hospital, Chiba, Japan
The pathogenesis of psoriatic itch is poorly understood. The aim of this study was to investigate the involvement of opioid receptors in scratching behaviour of imiquimod-induced psoriasis-like dermatitis model mice. Topical application of 5% imiquimod cream to the rostral back skin of mice induced antihistamine-resistant scratching behaviour. The expression of µ-opioid receptor (MOR) protein increased in the epidermis, dorsal root ganglia (DRG) and spinal cord of imiquimod-treated mice. In contrast, the expression of κ-opioid receptor (KOR) protein decreased in the DRG and spinal cord of imiquimod-treated mice, and was undetectable in the epidermis of both groups. Topical or intraperitoneal administration of the MOR antagonist naloxone and oral administration of the centrally acting KOR agonist ICI-199,441 inhibited scratching behaviour, whereas oral administration of the peri-pherally-selective KOR agonist asimadoline did not. These results suggest that peripheral and central MOR and central KOR may be involved in the modulation of scratching behaviour in imiquimod-treated mice.
Key words: imiquimod; itch; opioid receptor; psoriasis.
Accepted May 16, 2017; Epub ahead of print May 17, 2017
Acta Derm Venereol 2017; 97: xx–xx.
Corr: Kenji Takamori, Department of Dermatology, Juntendo University Urayasu Hospital, 2-1-1 Tomioka, Urayasu, Chiba 279-0021, Japan. E-mail: ktakamor@juntendo.ac.jp
Psoriasis is a common chronic inflammatory skin disease, and most patients experience itch (1). Such itch also reduces the quality of life (QOL) of patients with psoriasis (2).
Histamine is a major pruritogen, and histamine H1 receptor (H1R) antagonists are widely used to treat pruritus. H1R antagonists, however, are frequently ineffective in treating psoriatic itch (3, 4). Although other possible pruritogens and mediators may evoke itch sensation in psoriasis patients, none has been clearly demonstrated to cause itching (3). Thus, few therapeutic options are available to reduce itch, and it is important to develop new antipruritic treatments for patients with psoriasis.
Opioid receptors are classified into 4 types: µ-type (MOR), κ-type (KOR), δ-type (DOR) and nociceptin opioid (NOP)-type receptors (5, 6). These opioid receptors and their ligands are generally involved in pain sensation (7, 8). In particular, morphine and other MOR agonists are frequently used to treat pain associated with cancer (9). However, MOR agonists can cause pruritus as a side-effect (7, 10). Moreover, studies in primates and rodents have shown that MOR agonists evoke itch, whereas MOR antagonists suppress itch (11–16). In contrast, KOR agonists have been found to suppress itch in primates and rodents, whereas KOR antagonists have been shown to evoke itch in rodents (7, 17–20). We previously reported that imbalances in the expression and/or activation of the µ- and κ-opioid systems, rather than epidermal hyperinnervation, may be involved in the pathogenesis of psoriatic itch (21). Furthermore, a recent study reported that imbalance of epidermal MOR and KOR may be involved in the pathogenesis of psoriatic itch (22). Therefore, although the roles of opioid systems in the modulation of itch and pain sensations remain enigmatic (16, 23), these findings suggest that MOR and KOR may be promising therapeutic targets in patients with psoriatic itch.
This study pharmacologically investigated the roles of MOR and KOR in scratching behaviour in a mouse model of imiquimod (IMQ)-induced psoriasis-like dermatitis (24–26).
Male C57BL/6J mice (8–9 weeks old; Oriental BioService, Kyoto, Japan) were maintained in the experimental animal facility of Juntendo University Graduate School of Medicine under a 12-h light:12-h dark cycle at a regulated temperature of 22–24°C, with food and tap water provided ad libitum. Care and handling of all animals conformed to the NIH guidelines for animal research, and all animal procedures were approved by the Institutional Animal Care and Use Committee of Juntendo University Graduate School of Medicine.
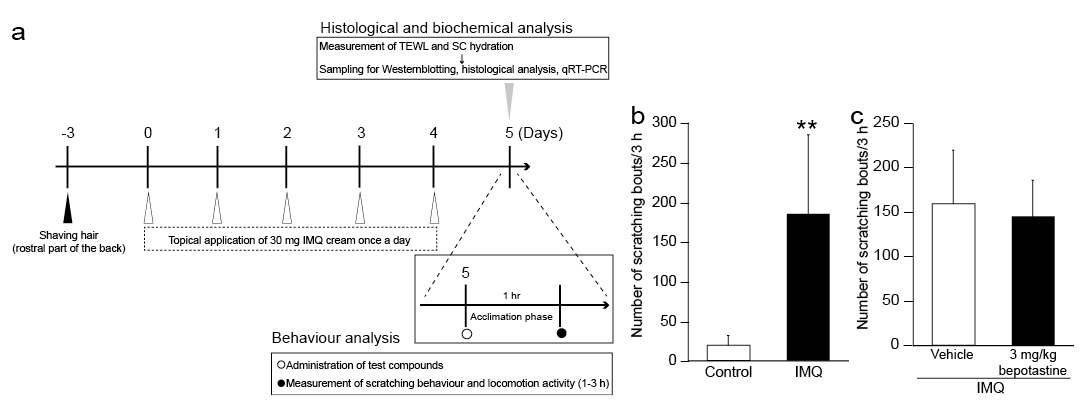
Mice were anaesthetized with sevoflurane, and the rostral part of the back was clipped with an electric shaver at least 3 days before the start of this study. Under sevoflurane anaesthesia, 30 mg commercially available 5% IMQ cream (Beselna Cream; Mochida Pharmaceutical, Tokyo, Japan) were topically applied to the shaved back skin each day for 5 consecutive days (Fig. 1a). Control mice were similarly shaved and anaesthetized with sevoflurane every day, but no IMQ or other creams were applied. Twenty-four hours after the 5th application of IMQ cream, scratching behaviour was monitored for 3 h using a SCLABA®-Real system (Noveltec, Kobe, Japan) (27).
Fig. 1. Spontaneous scratching behaviour in a mouse model of imiquimod (IMQ)-induced psoriasis-like dermatitis. (a) Treatment schedule. IMQ cream was applied topically to the rostral back skin of mice once daily for 5 days. Twenty-four hours after the 5th IMQ application, scratching behaviour was evaluated. (b) Number of scratching bouts over 3 h in control and IMQ mice. The number of scratching bouts was higher in the IMQ mice. (c) Number of scratching bouts over 3 h in mice treated with vehicle and 3 mg/kg bepotastine besilate (bepotastine). Bepotastine did not affect the scratching behaviour of the IMQ mice. Each bar in panels (b) and (c) represents the mean ± standard deviation (n= 8 per group). **p < 0.01 compared with the control group (unpaired t-test). SC: stratum corneum; TEWL: transepidermal water loss.
Bepotastine besilate (3 mg/kg), a histamine H1 receptor antagonist (Wako Pure Chemical Industries, Osaka, Japan); ICI-199,441 (1 mg/kg), a centrally acting KOR agonist (Santa Cruz Biotechnology, Dallas, TX, USA); asimadoline hydrochloride (10 mg/kg), a peripherally-selective KOR agonist (Sigma-Aldrich, St Louis, MO, USA), or vehicle (0.5% (w/v) sterilized methyl cellulose 400 solution; Wako Pure Chemical Industries) were administered orally (kg/10 ml) to the IMQ-treated mice. Naloxone (1 or 10 mg/kg), a MOR antagonist (Sigma-Aldrich) or vehicle (saline; Otsuka Pharmaceutical Factory, Tokushima, Japan) was injected intraperitoneally, and hydrophilic petrolatum (Maruishi Pharmaceutical, Osaka, Japan) containing 0.1 or 1% naloxone or vehicle (hydrophilic petrolatum) was applied topically (75 mg/mouse) to the IMQ-treated mice. One hour after oral or topical administration of test compounds, scratching behaviour and locomotion activity were monitored for 3 h using a SCLABA®-Real system and SCLABA®-Real tracking software (Noveltec). Locomotion activity, determined by total moving distance of centre of mass, was calculated from images recorded by SCLABA®-Real system. One hour after intraperitoneal administration of naloxone, scratching behaviour and locomotion activity were monitored for 1 h because naloxone has a short half-life in blood (28).
After sacrifice, dorsal skin, dorsal root ganglia (DRG) and spinal cord samples were excised from control and IMQ mice. Dorsal skin samples were treated with 0.25% trypsin/1 mM ethylenediaminetetraacetic acid (EDTA, Nacalai Tesque, Kyoto, Japan) overnight at 4°C to isolate the epidermis. Isolated epidermis, DRG and spinal cord were extracted with Radio-immunoprecipitation Assay (RIPA) buffer (Nacalai Tesque). Samples containing equal amounts of epidermis (20 µg), DRG (7.5 µg) and spinal cord (15 µg) proteins were applied to 15% Sodium Dodecyl Sulfate- Poly Acrylamide Gel Electrophoresis (SDS-PAGE) gels. The proteins were transferred onto Immobilon-P Transfer Membranes (Millipore) by Powered BLOT-One (ATTO Corporation, Tokyo, Japan). After blocking, the membranes were incubated with rabbit anti-MOR-1 antibody (1:1000 dilution; Santa Cruz Biotechnology) or goat anti-KOR-1 antibody (1:1000 dilution; Santa Cruz Biotechnology) overnight at 4°C. After washing, the membranes were incubated with horseradish peroxidase-conjugated anti-rabbit immunoglobulin G (IgG) (1:20,000 dilution; Jackson ImmunoResearch, PA, USA) or anti-goat IgG (1:20,000 dilution; Jackson ImmunoResearch). As a loading control, membranes were incubated with a horseradish peroxidase-conjugated mouse monoclonal antibody to glyceraldehyde-3-phosphate dehydrogenase (GAPDH; 1:10,000 dilution; Wako Pure Chemical Industries). Horseradish peroxidase-conjugated antibodies were detected using Chemi-Lumi One Ultra (Nacalai Tesque). The bands were subsequently quantified with ImageJ software (developed at the US National Institutes of Health; http://rsbweb.nih.gov/ij/).
Statistical analyses were performed using two-tailed Student’s t-tests and one-way analysis of variance with Dunnett’s multiple comparison test. All analyses were performed using Prism 6 software (GraphPad Software Inc., La Jolla, CA, USA), with p < 0.05 defined as statistically significant.
Psoriasis-like skin lesions were induced in the rostral back skin of C57BL/6J mice by topical application of IMQ once daily for 5 days (IMQ mice) (Fig. 1a and Fig. S1a, b). The number of scratching bouts was significantly higher in IMQ mice than in control mice (p = 0.0003) (Fig. 1b). In addition, dermatitis score, epidermal thickness and transepidermal water loss (TEWL) were significantly higher (p < 0.0001, p < 0.0001 and p < 0.0001), and SC hydration was significantly lower, in IMQ mice than in control mice (p < 0.0001) (Fig. S1c–f). Moreover, the levels of mRNAs encoding psoriasis-associated cytokines, such as interleukin (IL)-17A, IL-17F and IL-23p19, were significantly higher in the skin lesions of IMQ than of control mice (p = 0.0016, p = 0.0002 and p = 0.0001) (Fig. S1g–i).
We examined the effects of oral administration of the histamine H1R antagonist bepotastine besilate on scratching behaviour in IMQ mice (magnified view in Fig. 1a). Pretreatment with 3 mg/kg bepotastine besilate did not inhibit spontaneous scratching behaviour in IMQ mice (Fig. 1c). However, histamine-induced scratching behaviour was significantly reduced in mice treated with 3 mg/kg bepotastine besilate (p = 0.0021) (Fig. S2).
When we examined the effects of the topical MOR antagonist naloxone on scratching behaviour of IMQ mice, numbers of scratching bouts were significantly inhibited in mice treated with 0.1% and 1% naloxone than in mice treated with vehicle (p < 0.0001 and p < 0.0001) (Fig. 2a). There were no significant differences in locomotor activity among these 3 groups (Fig. 2b). Intraperitoneal administration of 1 and 10 mg/kg naloxone also resulted in a significant reduction in numbers of scratching bouts compared with vehicle-treated IMQ mice (p < 0.01 and p < 0.01) (Fig. 2c). The locomotor activity of mice treated with 10 mg/kg, but not 1 mg/kg, naloxone was significantly lower than that of mice treated with vehicle (p < 0.05) (Fig. 2d).
Fig. 2. Effect of topical and intraperitoneal naloxone on scratching behaviour and locomotor activity of imiquimod (IMQ) mice. (a) Number of scratching bouts and (b) locomotor activity over 3 h of IMQ mice pretreated with topical 0.1% or 1% naloxone or hydrophilic petrolatum (vehicle). Each bar represents the mean ± standard deviation (SD) of 8–16 mice per group. (c) Number of scratching bouts and (d) locomotor activity over 1 h of IMQ mice pretreated with intraperitoneal 1 or 10 mg/kg naloxone or saline (vehicle). Each bar represents the mean ± SD of 8–12 mice per group. *p < 0.05, **p < 0.01 compared with the vehicle group (one-way analysis of variance with Dunnett’s multiple comparison test).
Compared with vehicle, oral administration of 10 mg/kg asimadoline hydrochloride, a peripherally-selective KOR agonist, did not inhibit the scratching behaviour (Fig. 3a), and had no effect on locomotor activity of IMQ mice (Fig. 3b). In contrast, oral administration of 1 mg/kg ICI-199,441, a centrally acting κ-opioid receptor agonist, significantly reduced the number of scratching bouts of IMQ mice compared with vehicle (p < 0.0001) (Fig. 3c). There was no significant difference in locomotor activity between these 2 groups of mice (Fig. 3d).
Fig. 3. Effect of oral peripherally-selective and centrally acting κ-opioid receptor (KOR) agonists on scratching behaviour and locomotor activity of imiquimod (IMQ) mice. (a) Number of scratching bouts and (b) locomotor activity over 3 h of IMQ mice pretreated with oral 10 mg/kg asimadoline hydrochloride or vehicle. (c) Number of scratching bouts and (d) locomotor activity over 3 h of IMQ mice pretreated with oral 1 mg/kg ICI-199,441 or vehicle. Each bar represents the mean ± standard deviation of 8 mice per group. **p < 0.01 compared with the vehicle group (unpaired t-test).
Western blot analyses showed that MOR proteins were detected in the epidermis of IMQ, but not of control, mice (Fig. 4a). Expression of KOR proteins was undetectable in the epidermis of both groups (Fig. 4b). The level of expression of MOR proteins in the DRG of IMQ mice tended to be higher (Fig. 4c), whereas the level of expression of KOR proteins tended to be lower (Fig. 4d), than of control mice. Similarly, MOR expression in the spinal cords of IMQ mice was significantly higher (p = 0.0018) (Fig. 4e), while KOR expression tended to be lower (Fig. 4f), than of control mice.
Fig. 4. Expression patterns of µ-opioid receptor (MOR) and κ-opioid receptor (KOR) proteins in epidermis, dorsal root ganglia (DRG) and spinal cord of imiquimod (IMQ) mice. Expression of (a, c, e) MOR and (b, d, f) KOR proteins in (a, b) epidermis, (c, d) DRG and (e, f) spinal cord of control and IMQ-treated mice, as determined by Western blotting. The levels of expression of MOR and KOR in DRG and spinal cord were normalized to that of glyceraldehyde-3-phosphate dehydrogenase (GAPDH), with each bar representing the mean ± standard deviation of 4 samples per group. **p < 0.01 compared with the vehicle group (unpaired t-test).
The present study showed that IMQ mice displayed spontaneous itch-related scratching behaviour, as well as other clinical features of psoriasis-like dermatitis (Fig. 1b and Fig. S1). This scratching behaviour was not inhibited by oral administration of the H1R antagonist bepotastine besilate (Fig. 1c), at a dose (3 mg/kg) sufficient to inhibit histamine-evoked scratching behaviour (Fig. S2). These results suggest that a histamine-independent mechanism is primarily involved in the spontaneous itch-related behaviour of IMQ mice. This finding was consistent with the clinical features of psoriasis in humans (29), and a recent study involving IMQ mice (25).
Several pharmacological studies in primates and other animals have demonstrated that µ-opioid system plays a role in the induction of itch, whereas the κ-opioid system plays a role in the suppression of itch, at the central and/or peripheral levels (11, 30–32). In contrast, in chronic dry skin model mice, the numbers of scratching bouts of MOR and KOR knockout mice were almost same as of wild-type mice (23). Although the precise roles of these opioid receptors in the evocation of itch remains unclear, these results suggest that the balance of expression and/or activation between the MOR and KOR at least partly modulates itch sensation. Indeed, nalfurafine, a selective KOR agonist, has been effectively used to treat haemodialysis-associated pruritus (19) and pruritus of chronic liver diseases (33).
The present study found that the levels of MOR protein were higher in the epidermis, DRG and spinal cord of IMQ-treated than of control mice (Fig. 4a, c, e). In contrast, the levels of KOR protein tended to be lower in the DRG and spinal cord of IMQ than of control mice (Fig. 4d and f). In this study, we could not detect KOR expression in murine epidermis (Fig. 4b). Although possibly it is due to the sensitivity of Western blotting, we were also unable to detect KOR mRNA in murine skin by quantitative RT-PCR (Takahashi et al., unpublished observation). Both our study and that by Kupczyk et al. showed that MOR was unchanged and KOR was downregulated in the lesional skin of patients with psoriasis (21, 22). Although it is possible that expression patterns of epidermal MOR and KOR differ between humans and mice, these findings in IMQ mice suggest a relationship between opioid receptor imbalance at the central and/or peripheral levels and itch-related behaviour.
Our behavioural study found that intraperitoneal administration of the MOR antagonist naloxone significantly inhibited the number of scratching bouts in IMQ-treated mice (Fig. 2c). Central MOR was shown to be involved in morphine-induced scratching in mice (12). Western blotting of spinal cord proteins with an anti-MOR1 antibody that recognizes its N-terminus yielded double bands (Fig. 4e). The double bands might include MOR1D, one of the MOR1 isoforms involved in opioid-induced itch at the spinal cord (34). Although our study could not distinguish among individual MOR1 isoforms, these findings imply that central MOR, especially MOR1D, is involved in itch-related behaviour in IMQ mice.
Topical application of the MOR antagonist naltrexone has been shown to improve pruritus in patients with atopic dermatitis (14). Similarly, this study showed that topical naloxone treatment inhibited scratching behaviour in IMQ mice (Fig. 2a). Activation of the CB2 cannabinoid receptor was found to stimulate the release of β-endorphin, an endogenous MOR agonist, from rat epidermal keratinocytes, followed by the binding of β-endorphin to MOR on primary afferents, leading to anti-nociceptive effects (35). MOR is also expressed in sensory nerve fibres containing gastrin releasing peptide (GRP), an itch-related peptide, in murine skin (36). In addition, one study using MOR knockout mice found that MOR affected skin homeostasis (23). Taken together, these findings indicate that topical naloxone exerts anti-scratching effects in IMQ mice by modulating itch-related events, such as the production of certain pruritogens, and by relaying itch information from MOR-expressing cells and/or sensory nerve fibres at the periphery. In the present study, we found that anti-pruritic effects by topical application of naloxone were higher than by intraperitoneal injection. In contrast, it was reported that intraperitoneal injection of 0.5 mg/kg naloxone antagonized the analgesic effects of intrathecal morphine (37), and we showed the expression of MOR receptor proteins in the spinal cord of IMQ-treated mice was significantly higher than that of control mice. Based on these findings, it is considered that not only peripheral, but central, MOR were involved in the scratching behaviour of IMQ-treated mice, and peripheral MOR may play a more important role than central MOR in the scratching behaviour in this model mice.
The antipruritic effects of nalfurafine hydrochloride, a κ-opioid agonist, were shown to be mediated primarily by central KOR in humans and animals (18, 19, 38, 39). Pharmacologically, we found that oral administration of the centrally-acting KOR agonist ICI-199,441 significantly suppressed scratching behaviour in IMQ mice, whereas oral administration of the peripherally-selective KOR agonist asimadoline hydrochloride, at a dose (10 mg/kg) shown to have analgesic effects (40), did not inhibit scratching behaviour in IMQ mice (Fig. 3). In addition, we could not detect KOR in epidermis of control and IMQ mice (Fig 4a). These findings suggested that central, but not peripheral, KOR plays a pivotal role in spontaneous scratching behaviour in IMQ mice. Moreover, we found that scratching behaviour was associated with an imbalance between spinal MOR and KOR expression in IMQ mice (Fig. 4e and f).
A recent study reported that vascular endothelial growth factor-A (VEGF-A) is involved in the scratching behaviour of IMQ mice through epidermal hyperinnervation (26), whereas our immunohistological analyses showed that the number of intraepidermal nerve fibres was slightly, but not significantly, higher in IMQ than in control mice (Fig. S3), suggesting that epidermal hyperinnervation may be less involved in the spontaneous scratching behaviour of IMQ mice. Wong et al. (26) evaluated the total area revealed by fluorescence of PGP9.5-positive intraepidermal nerve fibres in IMQ-induced psoriasis-like dermatitis model mice, while we evaluated the number of PGP9.5-immunoreactive fibres within (Fig. S3b) and penetrating into the epidermis (Fig. S3c), in this model. Differences among the study results may be due to differences between their evaluation methods (area vs. number). Although involvement of intraepidermal nerve fibres in the scratching behaviour of IMQ mice was controversial, the results of our present study are supported by those of our previous study assessing the pathogenesis of itch in patients with psoriasis (21).
In conclusion, although the precise mechanisms underlying itch sensation controlled by MOR and KOR remain enigmatic, the results of the present study suggest that peripheral and central MOR and central KOR are involved in the modulation of spontaneous itch-related behaviour in IMQ-treated mice.
This work was supported by a grant of Strategic Research Foundation Grant-aided Project for Private Universities from MEXT (S1311011).
The authors declare no conflicts of interest.


 Click to show fullsize
Click to show fullsize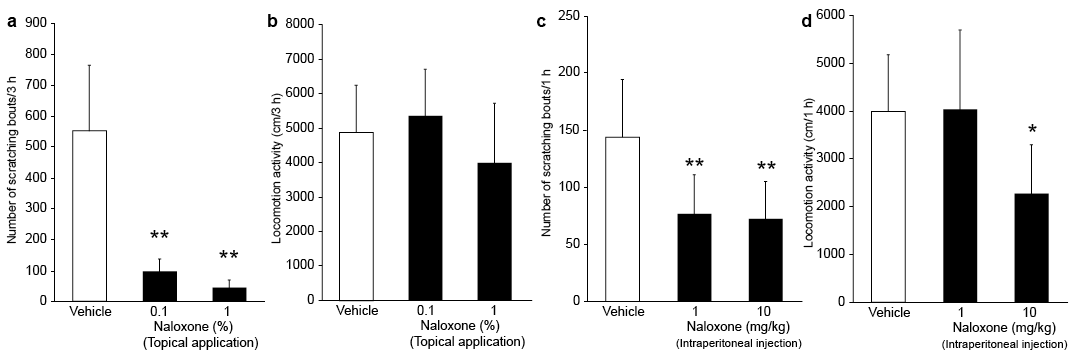 Click to show fullsize
Click to show fullsize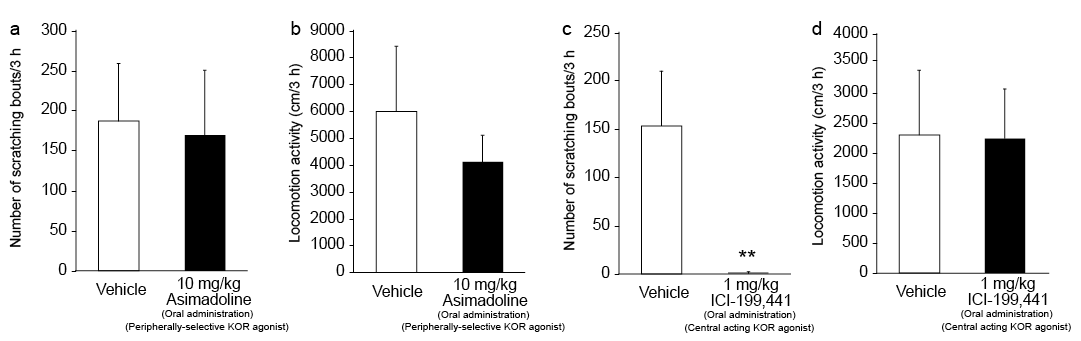 Click to show fullsize
Click to show fullsize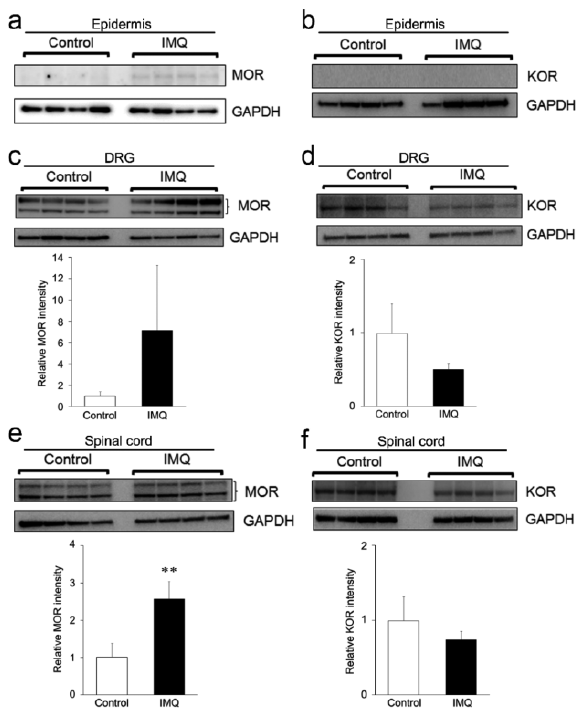 Click to show fullsize
Click to show fullsize