


1Laboratory for Experimental Cutaneous Pain and Itch Research, SMI®, Department of Health Science and Technology, Faculty of Medicine, Aalborg University, Fredrik Bajers Vej 7, Bld. A, A2-208, DK-9220 Aalborg, and 2The Allergy Clinic, Department of Dermato-Allergology, Copenhagen University Hospital, Gentofte, Copenhagen, Denmark. *E-mail: hha@hst.aau.dk
Accepted Oct 15, 2018; Epub ahead of print Oct 15, 2018
Itch is a frequent symptom in many dermatological, systemic and neuropathic diseases (1). Several populations of thin primary afferents respond to a variety of pruritogens (2). In many diseases, such as atopic dermatitis (AD), itch is thought to be transmitted non-histaminergically (3). This fits the observation that antihistamines are ineffective in AD (4). For mechanistic studies, pharmaceutical proof-of-concept studies, and in relation to sensory phenotyping of chronic itch patients, reliable provocation models of histamine-independent itch are important (5). The cowhage-model has become the “gold standard” of non-histaminergic evoked itch in humans (6, 7), and while it is a very effective model it suffers from several drawbacks: (i) carefully controlled applications of the spiculae are impossible, (ii) no commercial outlet supplies the spiculae, (iii), and batch-to-batch and inter-strain differences appear to be present. With this in mind, we aimed to develop a new and easily standardizable itch model relying on β-alanine, a naturally occurring amino acid. β-alanine evokes itch and a pricking sensation upon intradermal injection through activation of superficially terminating mas-related G-protein-coupled receptor (Mrgpr) D-expressing neurones (8, 9). In non-human primates the primary afferent C-fibre population activated by β-alanine is distinct from that activated by histamine (10). The present study characterized the dose-response features of the β-alaninergic model of itch introduced with skin prick test (SPT) lancets in human skin. For comparative purposes itch provocations with cowhage and histamine were conducted and hyperknesis and cutaneous flare reactions were monitored.
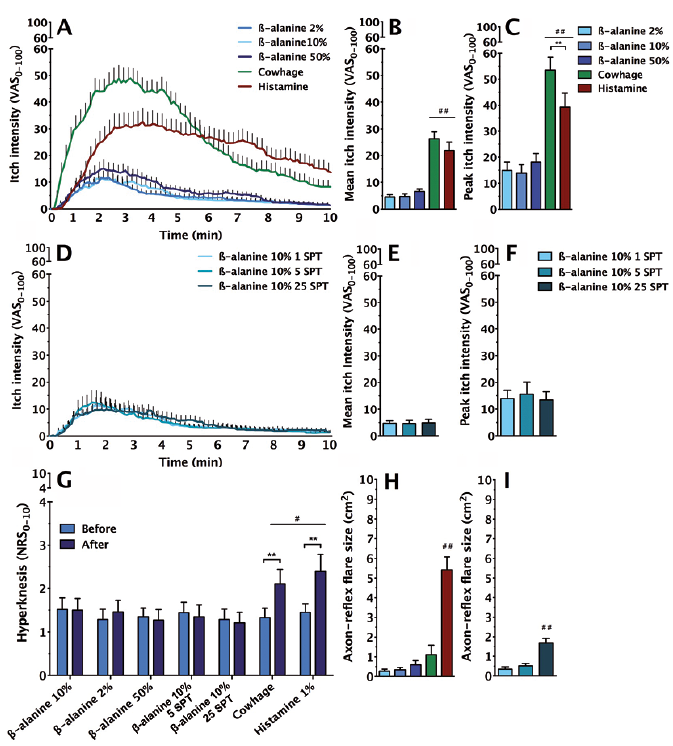
β-alanine introduced with SPT lancets did not exhibit clear dose-response features (no differences between any of the β-alanine conditions) (Fig. 1A–C). This was true both for the mean and peak itch intensities and also independent of the numbers of SPT lancet punctures conducted, i.e. 1 vs. 5 vs. 25 (Fig. 1D–F). Compared with both histamine and cowhage, β-alanine induced significantly less robust itch. The temporal profiles of itch were different between cowhage and histamine, with cowhage exhibiting a more rapidly occurring peak and a faster decline than histamine. The temporal profile of β-alanine-induced itch resembled a scaled down version of cowhage-evoked itch.
Fig. 1. (A) Temporal profile, (B) mean, and (C) peak itch intensities induced by different pruritogens. Cowhage and histamine induced significantly higher peak and mean itch than all other provocations. Temporal profile (D) as well as (E) mean and (F) peak itch intensities induced by 10% β-alanine when administered by 1, 5 or 25 skin punctures. No differences were observed. (G) Hyperknesis was present only following cowhage and histamine provocations. (H) Only histamine produced a robust axon-reflex-flare, while the 25 skin prick procedure caused (I) a small, but significant, flare reaction. NRS: numerical rating scale; SPT: skin prick test administration (i.e. number of punctures); VAS: visual analogue scale. Note: B, C and H as well as E, F and I share legends. *p < 0.05, **p < 0.01, #/##significant from all other conditions at p < 0.05 or p < 0.01, respectively.
β-alanine never induced significant mechanical hyper-knesis (Fig. 1G). In contrast, cowhage and histamine produced mechanical hyperknesis of similar intensity (approximately 60% increases in mechanical itch sensitivity).
Akin to cowhage, β-alanine did not produce an axon-reflex flare, regardless of the applied concentration (Fig. 1H), but a small homotopic erythema of 1.7 cm2 was recorded in the 25-punctures condition (Fig. 1I). As expected, histamine produced a pronounced neurogenic inflammatory response and a circumscribed wheal reaction.
The doses of β-alanine used evoked only mild itch in comparison with cowhage and histamine. Furthermore, stimulus-response effects were non-existent, both in the range 2–50% and when changing the number of SPT punctures. β-alanine has previously been shown to induce mild itch and pricking on intradermal injection assessed by a labelled magnitude scale (8). The relative relationship between β-alanine and cowhage-evoked itch presently observed corresponds approximately to previous data irrespective of the psychometric scale applied (cowhage evoking approximately 1.5–3 times more intense itch than β-alanine). For intradermal injections, concentrations of 50–200 mM β-alanine produced ambiguous or limited dose-response effects (8). Finally, rodent data confirm β-alanine as a weak pruritogen generally evoking 2–4-folds less scratching than other commonly used pruritogens, such as histamine and chloroquine (14).
As opposed to cowhage and histamine, β-alanine did not establish significant secondary mechanical hyperknesis. Secondary hyperknesis is thought to reflect sensitization of spinothalamic tract neurones transmitting itch in the spinal cord following robust peripheral pruriceptive barrage (2, 13). Given the lack of robust evoked itch in the β-alanine conditions, it is thus not surprising that no significant hyper-knesis was observed (13).
A small homotopic erythematous reaction was observed following the administration of β-alanine using 25 skin punctures, probably evoked by a superficial microtrauma caused by repeated pinpricks. A discernible axon-reflex flare was observed only in the histamine condition. This likely reflects that only histamine effectively activates mechano-insensitive C-fibres, capable of producing a pronounced flare reaction (15), while cowhage and β-alanine act on mechano-heat C-fibres expressing PAR-2/ MrgprX1/2 (cowhage) and MrgprD (β-alanine).
Two limitations must be kept in mind: (i) the lack of a dose effect could be a consequence of applying concentrations all at the upper plateau of the stimulus-response curve. However, with a 2–50% range including assessment of the influence of numbers of SPT punctures, this seems unlikely. (ii) The biochemical properties of β-alanine could make it ill-suited for punctate delivery and prevent it from reaching its substrate nerve-endings. This also appears unfeasible given the known superficial termination of MrgprD-expressing C-fibres (9), and the lack of an effect of multiple SPT punctures (16).
While β-alanine is an easily standardizable human provocation model capable of evoking mild itch by both punctate delivery and intradermal injection, it is much less robust than the gold standard models utilizing histamine and cowhage. However, it could perhaps be applied in study designs where mild and brief itch is desired. Future studies in humans should assess other purported histamine-independent itch mediators than β-alanine, in order to successfully establish a reliable non-histaminergic itch model alternative to cowhage.


 Click to show fullsize
Click to show fullsize