


1Department of Psychology, University of Hull, Hull, and 2Birmingham City University, Faculty of Health, Education and Life Sciences, Birmingham, UK
While temperatures in the noxious range are well-known to inhibit acute itch, the impact of temperature in the innocuous temperature range is less well understood. We investigated the effect of alternating short-term temperature changes in the innocuous range on histamine and cowhage-induced acute itch, taking into account individual differences in baseline skin temperature and sensory thresholds. Results indicate that cooling the skin to the cold threshold causes a temporary increase in the intensity of histamine-induced itch, in line with previous findings. Skin warming increased cowhage-induced itch intensity. Potential mecha-nisms of this interaction between thermosensation and pruritoception could involve cold-sensitive channels such as TRPM8, TREK-1 or TRPC5 in the case of histamine. The rapid modulation of cowhage induced itch – but not histamine-induced itch – by transient skin warming could be related to the lower temperature threshold of pruriceptive polymodal C-fibres (cowhage) as compared to the higher temperature
threshold of the mechanoinsensitive C-fibres conveying histaminergic itch.
Key words: skin temperature; histamine; sensory thresholds; pruritus.
Accepted Oct 25, 2018; Epub ahead of print Oct 25, 2018
Acta Derm Venereol
Corr: Dr Henning Holle, Department of Psychology, University of Hull, Cottingham Road, Hull HU6 7RX, U.K. E-mail: h.holle@hull.ac.uk
Determining the influence of temperature changes in the non-painful range on itch is not only of clinical interest, but also relevant for basic psychophysical research, which often requires that itch intensity can be reliably modulated within a matter of seconds in an on–off fashion. We characterized the effect of short-term temperature changes on two types of itch sensations; histamine and cowhage-induced itch. Our result shows that both itch pathways can be modulated by short-term changes in temperature, potentially enabling statistically powerful neuroimaging studies, to further elucidate the cortical network underpinning the clinically relevant sensation of itch.
It is well documented that thermal counter-stimulation in the noxious range reduces the intensity of acute itch (1–4). However, the effect of brief, less extreme changes of temperature on itch, either by slightly increasing or decreasing the skin temperature from its baseline temperature of about 32°C, is less well understood. Determining this relationship is not only of clinical interest, but also relevant for basic research on itch in humans, which often requires an experimental itch model where itch intensity can be quickly modulated (5).
Several studies have investigated the effect of innocuous warmth on experimentally induced acute itch. Ward et al. (2) induced itch using histamine iontophoresis and found that warming the skin directly adjacent to the itch induction site to 38°C using a thermode did not influence itch intensity. Yosipovitch et al. (4) also used histamine iontophoresis in combination with a thermode applied 3 cm distal to the itch induction site, and again found that repeatedly warming the skin to 41°C did not modulate itch intensity. Fruhstorfer and colleagues (1) used a slightly different approach in that the thermode was placed directly on the itching skin site. Skin temperature was then slowly increased from baseline at a rate of 0.5°C/s until heat pain was reported. They observed large inter-individual differences in the effect of warmth. The majority of participants (n = 23) reported a decrease in itch intensity as temperature increased. Notably this effect was not restricted to the noxious range, but also evident at sub-noxious temperatures below 40°C. A smaller subset of participants (n = 7) reported the opposite pattern, with itch increasing as temperature increased. A final subset (n = 4) showed no influence of warmth on itch.
In summary, findings about a potential effect of innocuous warmth on itch are mixed. The evidence suggests that directly warming the affected skin site (1), as opposed to an adjacent skin site (2, 4), may be a more sensitive approach. Another limitation of existing studies is that inter-individual differences in baseline skin temperatures and individual warm thresholds have so far not been taken into account (6).
Regarding the effects of cooling on acute itch, findings are, at first glance, contradictory. Yosipovitch et al. (4) observed that repeatedly cooling skin adjacent to the itch site to 15°C (i.e., at the threshold from innocuous to noxious cold) does not influence itch intensity. Fruhstorfer and colleagues (1) observed that slowly cooling the skin, from baseline at a rate 0.5°C/s until cold pain was perceived, led to a decrease in itch intensity. This effect was consistent across subjects and began to manifest itself already in the innocuous temperature range below 25°C. Andersen et al. (7) observed significant reductions of itch intensity when skin was cooled to temperatures of 22°C, 12°C and 4°C. In contrast, Pfab et al. (5) observed that fast, short-term cooling of the skin, with temperature alternating between 32°C and 25°C every 20 s, led to a significant increase in itch intensity during cool periods. These opposing effects of skin cooling on histaminergic itch – with long-lasting cooling having an inhibitory, but short-term phasic cooling an excitatory influence on acute itch – may be related to adaptation effects. Unlike receptors in the noxious temperature range, innocuous thermoreceptors are fast adapting (8). The Transient receptor potential cation channel subfamily M member 8 (TRPM8), a major thermoreceptor for innocuous cold, shows a rapid increase in firing rate upon moderate cooling, but also rapid adaptation within the first 8 s of a cold ramp (9).
The present study aims to extend existing knowledge in several ways. First, we will explore the effects of rapid cooling and warming of the skin on histamine-induced itch, taking into account individual differences in baseline skin temperature and individual warm and cold thresholds as determined via quantitative sensory testing. Second, we will also characterize, for the first time, the influence of temperature on non-histaminergic itch. The cowhage provocation paradigm has recently been established as an alternative experimental itch model (10) and is taken to be more characteristic of itch occurring in chronic pruritic diseases such as atopic dermatitis (11). Investigating the potential modulatory influence of innocuous temperature changes is not only potentially of clinical benefit, but also important for basic psychophysiological studies, which often require that itch intensity can be modulated within a matter of seconds (5, 12).
All participants were right-handed students of the University of Hull. Exclusion criteria included a history of allergy, acute or chronic skin conditions, vascular disease, low blood pressure, asthma, histamine intolerance, sensitive skin, hypersensitivity to certain food types and any intake of drugs. The full exclusion criteria are detailed in the protocol approved by the University of Hull ethics committee in accordance with the Declaration of Helsinki. All participants were naïve as to the purposes of the experiment and gave full written informed consent. In Experiment 1, 17 healthy volunteers (14 females), aged between 18 and 34 years (mean age 21.8) took part. Fourteen healthy volunteers (11 females), aged 18 to 35 years (mean age 21) were recruited for Experiment 2. Data from one participant was excluded due to failure to comply with the itch rating procedure in Experiment 2.
In Experiment 1, itch was induced using the histamine skin prick test. One drop of 1% histamine dihydrochloride in aqueous solution was applied to the volar aspect of the forearm and then the skin superficially punctured with a special lancet (Allergy Therapeutics, Worthing, UK). Care was taken to ensure epidermal histamine delivery. Had any bleeding occurred the test session would have been halted and rescheduled. All skin pricks were done by the same female investigator to minimize variability in the application technique. After an interval of at least 5 min, during which the participant reported no further itching, a second trial of itch stimulation was performed on the alternate arm. Stimulation side was counterbalanced across participants.
In Experiment 2, histamine-independent itch was induced using cowhage spicules. Mucunain, the itch-inducing agent of the cowhage-spicules, has previously been shown to activate Proteinase Activated Receptors 2/4 (PAR 2/4) in the epidermis (13), but recent evidence suggests that cowhage-evoked itch may in fact in part or wholly be mediated by members of the family of Mas-related G-protein coupled receptors (MRGPRX1 and MRGPRX2 14). Cowhage induces itch that differs from histamine-evoked itch in terms of its local skin reaction (little or no flare for cowhage (15, 16), the type of nociceptive C fibres involved (cowhage: mechanosensitive; histamine: mechanoinsensitive (17, 18), and the quality of the itch response (cowhage: more pricking, stinging and burning (10, 16, 19)). Forty cowhage spicules were counted under a magnifying glass, picked-up by microtweezer and applied within a 4 cm2 area on the skin. The spicules were gently rubbed for 45 s onto the volar aspect of the forearm, 3 cm proximal of the distal wrist crease, with a circular motion to facilitate contact. Scotch Tape (3M, St Paul, MN) was used to demarcate the area of the forearm to prevent any stray spicules from stimulating surrounding skin. Cowhage spicules were removed at the end of the rating period using Scotch Tape.
Each experimental session started with a familiarization trial of the itch stimulus and rating scale, as recommend by Phan et al. (20), followed by determination of individual warm and cold thresholds (WT and CT, respectively), and two temperature trials. During these temperature trials, each lasting 540 s, an initial itch stimulus was either accompanied by temperature alternating between baseline and WT (warm trial) or temperature alternating between baseline and CT (cold trial). The order of warm and cold trials was counterbalanced across participants.
In Experiment 1 only, we additionally assessed the qualitative experience of itch using the modified version of the Eppendorf Itch Questionnaire (EIQ, 19) and the size of the skin reaction (wheal and flare).
Starting with the histamine prick, participants rated itch intensity using the general version of the Labelled Magnitude Scale (gLMS) (21, 22). The gLMS consists of a vertical line presented on a computer screen with quasi-logarithmically placed labels of “no sensation” at 0, “barely detectable” at 1, “weak” at 6, “moderate” at 17, “strong” at 35, “very strong” at 53 and “strongest imaginable sensation” at 100. Participants were asked to continuously rate the itch sensation experienced for 540 s beginning immediately following itch application, by using the scroll wheel on a computer mouse to move a red line up or down the vertical line on the screen to indicate the required rating. Ratings were sampled at a frequency of 1 Hz. The mouse was operated face down using the non-stimulated hand as both forearms were in a palm up position throughout all conditions.
After the familiarization, baseline skin temperature of the stimulation area was taken, followed by a determination of individual warm and cold thresholds (WT and CT) using the method of limits (Experiment 1: Mean baseline ± standard deviation (SD): 31.2 ± 0.83°C, CT = 25.3 ± 1.03°C, WT = 34.7 ± 0.99°C; Experiment 2: Mean baseline ± SD: 31.42 ± 1.09°C, CT = 24.62 ± 0.89°C, WT = 35.65 ± 1.19°C). Briefly, determining WT and CT involved placing a Medoc Pathway Cheps (27 mm diameter) thermode on the volar aspect of the forearm, 3 cm proximal to the crease of the wrist. Baseline temperature was set at 32°C. The mean threshold temperature of 5 consecutive measurements was calculated, starting with determination of WT. After a 5 min break, the CT was determined in the same fashion. All thresholds were obtained with ramped stimuli (1°C/s), and ramps always started at the baseline temperature. Participants were instructed to verbally respond once they felt their skin becoming either warm or cold. Note that this mode of instruction is different from the instruction given to participants in other studies (e.g., “Press the button as soon as you feel the slightest change of temperature”, 6) which may explain why WT and CT values in our studies are more extreme (lower CTs and higher WTs) than in previous studies (6, 23–25).
After determination of CT and WT, the two experimental trials were presented (i.e., a warm trial and a cold trial). In the each of these trials, itch was induced on the dorsal forearm as described above. Immediately following itch application, skin temperature was modulated using the thermode placed directly onto the test area. Each trial consisted of a number of cycles of with a length of 40 s each (Experiment 1: 13 cycles, Experiment 2: 12 cycles). Each cycle began with a neutral block (at baseline temperature) followed by either a cold block where skin temperature was at that individual’s CT or a warm block (skin temperature at WT) depending on the condition. Each block lasted 20 s, with temperature change occurring at the end of each block at a rate of 5°C/s. In Experiment 1, the first temperature change occurred 20 s after itch application (see also Fig. 1). In Experiment 2, where cowhage spicules were applied during a 40 s initial rub-in period, the first stimulation cycle was initiated 60 s after itch application compared with 20 s in Experiment 1 resulting in 12 equal cycles.
The analysis of continuous computerised data was simplified by using the itch intensity rating at the mid-point of each 20 second block, a point in time by which the target temperature was always fully established. In Experiment 1, this provided 13 ratings each for neutral and cold in the innocuous cold trial and 13 ratings for neutral and warm each in the innocuous warmth trial. In Experiment 2, the number of ratings used was 12, respectively.
Two complementary statistical analyses were performed. In a first analysis, we directly compared the cold and warm condition with each other. The Itch-Only condition was not included as a full experimental condition, but to familiarize participants with the experience of experimentally induced itch and usage of the rating scale. Therefore, the Itch-Only condition was always presented first and not included in the counterbalancing procedure. For the same reason, we cannot perform meaningful statistical comparisons against the Itch-Only condition. In a second analysis, we asked whether the transient temperature change within each time course (from baseline to cold, or from baseline to warm) led to a systematic change in itch intensity. In line with previous studies on the effect of transient temperature on itch (4, 5, 26), this second analysis focused on the effect of temperature after itch had become fully established. Thus, the analysis was restricted to the descending flank of the itch response (i.e., in Experiment 1 the first 100 s of itch ratings were excluded, see Fig. 1, in Experiment 2 the first 140 s were excluded, see Fig. 3).
To analyse the transient effect of temperature on itch, psycho-physical ratings of itch were analysed using a two-factor (Time and Temperature) repeated measures ANOVA to determine transient changes in itch sensation due to change in temperature and the passage of time (for a similar approach, see 4). There were two levels of the Temperature factor (baseline vs innocuous warmth or baseline vs. innocuous cold) and several levels of the Time factor (Experiment 1: 11 levels, Experiment 2: 10 levels). A significant main effect of either the Temperature factor or a significant interaction between the Temperature and Time factors indicate that the perception of itch is significantly influenced by the presence of the temperature stimulus. Greenhouse & Geisser (27) correction was applied where necessary.
In both experiments, standardized residuals of the itch ratings significantly deviated from a normal distribution (Experiment 1: all Shapiro-Wilk’s W(17) < 0.834, all p < 0.006; Experiment 2: Shapiro-Wilk’s W(12) = 0.849, p = 0.035). We therefore excluded those participants with the largest residuals from the statistical analysis (3 participants in Experiment 1, 2 participants in Experiment 2), resulting in a normally distributed sample (Experiment 1: n = 14; Experiment 2: n = 11) for analysis of the itch rating data. The exclusion of these participants does not change the pattern of significant effects in the analyses described below.
After each trial in Experiment 1, a shortened version of the EIQ (19), adapted for qualitative and quantitative assessment of pruritus in healthy volunteers, was completed by each participant. The 23 statements are designed to measure and identify different sensory qualities of the itch sensation. Each statement is rated on a 5-point scale from 0 (not appropriate) to 4 (absolutely appropriate). The modified EIQ was not administered in Experiment 2.
In Experiment 1, at the end of each itch trial, but before completion of the EIQ, the magnitude of the histamine-induced skin reaction was evaluated by measuring the size of the wheal (vascular leakage response to histamine, observed as a raised, often pale and circumscribed dermal oedema, 28) and flare (increased superficial perfusion following a histamine prick, 28). Wheal and flare response from the application of histamine were photographed, alongside a measurement scale for post-test analysis. This analysis was undertaken by a third party who was blind to the experimental condition to which the photograph pertained. The wheal and flare area were identified using a Graphical Editing Software (www.gimp.org) and converted to cm2 using the scale contained in each photograph. Wheal and flare photographs from three participants were missing. Since there is usually no wheal and only a minimal flare in reaction to cowhage-induced itch (10, 19), no skin reaction measurements were taken in Experiment 2.
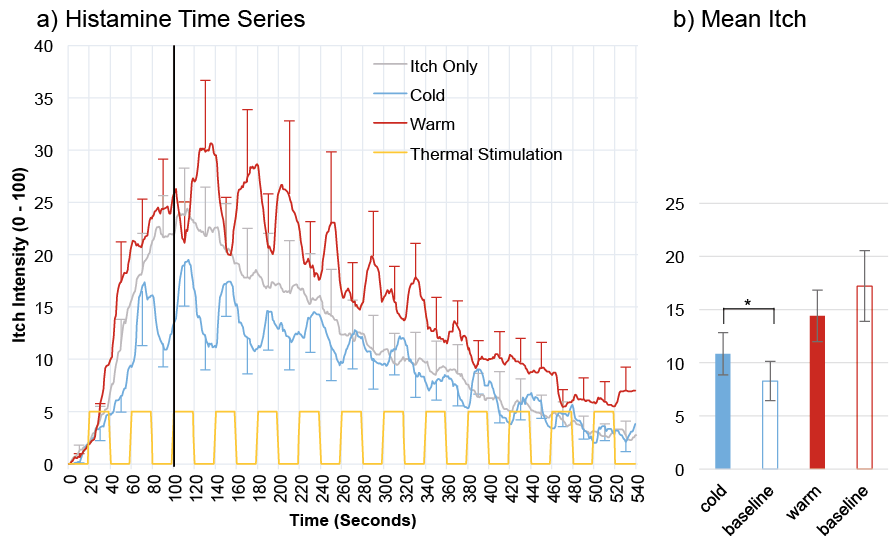
As can be seen in Fig. 1, histamine elicited the typical itch response, peaking about 100 s after the onset of stimulation, followed by a slow decay. Regarding the overall effect across the entire time series, skin warming seems to lead to a long-lasting increase in itch intensity, relative to skin cooling. When looking at the transient effect of temperature change (i.e., 20 s periods of temperature change alternating with 20 s of baseline), there is an apparent trend that warming the skin temporarily decreases itch, followed by a rebound when temperature returns to baseline levels. Cooling the skin seems to have the opposite transient effect on histamine-induced itch, with an increase in itch observed during cool periods, followed by increase when temperature has returned to baseline.
Fig. 1. a) Mean itch intensity ratings (n = 14) over 540 s for innocuous cold, innocuous warmth and the familiarisation procedure (Itch-Only). The pattern of thermal modulation for 13 temperature cycles is shown at the bottom. Time 0 represents the onset of the histamine prick. The thick vertical represents the beginning of the segments used for statistical analysis of the transient effect. The standard error of the mean is provided for the mid-cycle ratings that were entered into the ANOVA (cycles 3 to 13). b) Illustration of the main effect of Temperature (altered temperature vs. baseline, as determined in the ANOVA), separately for cold and warm conditions. The Itch-Only condition was always presented first and was not included into the counterbalancing procedure (see Methods section for underlying rationale). *Significant effects (p < 0.05.
Histamine: Overall effect of warm vs. cold. Itch ratings were significantly (t(13) = 2.873, p = 0.013) higher during the warm condition (mean 15.39, SE = 2.36), relative to the cold condition (mean 9.19, SE = 2.36). This difference between warm and cold trials was robust both during periods of temperature stimulation (t(13) = 2.158, p = 0.050) as well during intermittent baseline periods (t(13) = 2.475, p = 0.028).
Histamine: Transient effect of temperature change vs. baseline. The ANOVA for the cold condition showed a significant effect of temperature, (F (1, 13) = 5.442, p = 0.036), indicating that cooling the skin elicited higher itch ratings (mean 10.825, SE = 1.988) relative to when skin temperature was at baseline (mean 8.279, SE = 1.861). There was also a significant main effect of time, (F (1.765, 22.939) = 7.727, p = 0.004), but no significant interaction (F (2.696, 35.054) = 1.063, p = 0.372) suggesting that the itch enhancing effect of transient cooling of the skin is maintained throughout the descending flank of the itch response.
In contrast, the ANOVA for the warm condition only revealed a significant main effect of time (F (1.612, 20.958) = 8.675, p = 0.003) but no main effect of temperature (F (1, 13) = 0.795, p = 0.389) and no significant interaction, (F (1.491, 19.380) = 0.851, p = 0.412) suggesting that a short-term increase in temperature does not systematically affect subjective itch ratings. Visual inspection of single subject time-courses indicated that only a subset of participants (n = 4) reported strong experiences of itch relief when skin was warmed to the WT (which is also reflected in the mean response shown in Fig. 1), but this response pattern was not sufficiently systematic across participants to reach statistical significance.
Histamine: Effects of temperature on sensory quality of itch sensation. The qualitative experience of the histamine prick test, as indicated by the abbreviated EIQ, can be seen in Fig. 2. Since standardized residuals were not normally distributed, the differences between warm and cold conditions for each of the 23 questionnaire items were evaluated using Wilcoxon signed rank tests. These tests indicated that relative to the cold condition, the itch during warm trials was perceived as more sharp (p = 0.021), pulsating (p = 0.037), stinging (p = 0.008), warm (p = 0.019) and burning (p = 0.022). Note that these tests were of an exploratory character and therefore not corrected for multiple comparisons.
Fig. 2. Mean rating for items of the adapted-Eppendorf Itch Questionnaire (19) following histamine-induced itch. Rating scale ranged from 0 (not appropriate) to 4 (absolutely appropriate). *Items were ratings significantly differed (p < 0.05) between the cold and warm condition.
Histamine: skin reaction. We also analyzed whether thermal stimulation affected the size of the skin reaction typically elicited by a histamine prick, i.e. wheal and flare size (see Table I). The corresponding paired t-tests indicated that cooling the skin, relative to warming, significantly reduced the flare size (t(13) = 2.23, p = 0.044). Wheal size was not affected by temperature (t(13) = 1.01, p = 0.332).
Table I. Wheal and flare size for experimental conditions in cm2 (n = 14)
As can be seen in Fig. 3, cowhage elicited a typical itch response, peaking around 140 s after the onset of itch induction. There does not seem to be an overall difference between the warm and cold conditions for cowhage-induced itch. However, there appears to be a transient effect of temperature within the warm condition, with itch intensity increasing whenever the skin temperature was raised to the warm threshold, followed by a reduction when skin temperature returned to baseline level.
Fig. 3. Mean itch intensity ratings (n = 11) of cowhage-induced itch for the familiarization session (Itch Only) and the two experimental trials. Time 0 represents the beginning of itch induction. Only ratings obtained during the descending flank of the itch response (i.e., to the right of the thick black vertical line) were included in the statistical analysis. b) Illustration of the main effect of Temperature (altered temperature vs. baseline, as determined in the ANOVA), separately for cold and warm conditions. The Itch-Only condition was always presented first and was not included into the counterbalancing procedure (see Methods section for underlying rationale). *Significant effects (p < 0.05).
Cowhage: Overall effect of warm vs. cold. There was no significant overall difference between the warm and the cold condition for cowhage induced itch, t(9) = –0.272, p = 0.792. There were also no significant differences when this comparison was made separately for periods of temperature stimulation (t(9) = –0.827, p = 0.430) and intermittent baseline periods (t(9) = 0.943, p = 0.370).
Cowhage: Transient effect of temperature. The ANOVA for the warm trials revealed a significant main effect of temperature (F(1, 10) = 6.337, p = 0.031), indicating that transient warming of the skin significantly enhanced cowhage-induced itch (mean 13.527, SE = 3.156) relative to when skin temperature was at baseline (mean 8.814, SE = 2.149). Additionally, there was a significant main effect of time (F(1.35, 13.50) = 11.03, p = 0.003). The interaction was not significant (F(2.68, 26.78) = 3.029, p = 0.052).
In contrast, the ANOVA for the cold trials only yielded a significant main effect of time (F (1.43, 12.87) = 12.01, p = 0.002). The main effect of temperature (F (1, 9) = 0.93, p = 0.360) as well as the interaction between time and temperature (F (2.08, 18.69) = 0.94, p = 0.410) were not significant, suggesting a transient reduction in temperature does not have a systematic effect on cowhage-induced itch.
The aim of the present study was to investigate the effect of alternating short-term temperature changes in the innocuous temperature range on histamine and cowhage-induced acute itch, taking into account individual differences in baseline skin temperature and sensory thresholds. Results suggest that repeatedly cooling the skin to the cold threshold causes a small but statistically reliable temporary phasic increase in the intensity of histamine-induced itch, in line with previous findings. Transient warming of the skin did not consistently influence histaminergic itch. An opposite pattern was observed for cowhage-induced itch, where transient cooling had no systematic effect, but repeated cycles of warming led to phasic increases in itch intensity.
A novel finding of the present study is that we demonstrate, for the first time, the effect of innocuous temperature change on cowhage-induced itch. We found that a short-term increase of the skin temperature to the individual warm threshold is accompanied by an increase in the intensity of cowhage-induced itch. This raises the possibility that itch intensity can be rapidly modulated, within a matter of seconds, which is a requirement for many psychophysiological studies, including functional magnetic resonance imaging. Existing studies on the cortical network underpinning cowhage-induced itch are sparse (29). The findings of the present study raise the possibility that a statistically powerful paradigm involving temperature modulation of itch intensity, as it has been developed for histaminergic itch (5), can now be developed for cowhage as well. However, one limitation that first needs to be addressed is the currently somewhat limited effect size of the temperature effect for cowhage. One way to enhance the effect size for this purpose could be to switch from continuous to time-bound independent itch ratings (e.g., only one rating per temperature cycle, see, for example, 26).
Our finding that short-term cooling elicits a temporary increase in histamine-induced itch is in line with previous findings by Pfab and colleagues (5). They observed that repeatedly cooling the skin from 32°C to 25°C reliably modulates itch, a finding that has since been replicated several times (12, 30–32). Our results show that the same effect can also be elicited when the skin is cooled down to the individual cold threshold.
A potential mechanism for the cooling effect on histaminergic itch could involve the TRPM8 receptor (33–35), the main thermoreceptor for innocuous cold (9). Interestingly, continuous chemical activation of this receptor using menthol reduces both acute histaminergic (36) as well as chronic itch (37), whereas transient activation via phasic skin cooling increases acute histaminergic itch (the present study, 5, 26). As has been mentioned in the introduction, a possible explanation of this seemingly paradoxical effect of skin cooling on histaminergic itch (long-lasting cooling decreases itch, where short-term phasic cooling increases it) are adaptation effects. The TRPM8 receptor shows a rapid increase in firing rate upon moderate cooling, followed by rapid adaption within the first 8 s of a cold ramp (9). This temporal profile is in line with psychophysical and neuroimaging studies reporting a steep increase in histamine evoked itch intensity (5) and brain activity in itch-associated brain areas (12) during the first 8 s of skin cooling. Thus, in studies applying a continuous moderate cooling of the skin, the TRPM8 receptors are mostly in a state of adaptation. In contrast, repeated phasic cooling of the skin results in more repeated bursts of TRPM8 activity at the beginning of each cold ramp, which may lead to a phasic increase in itch. However, a problem for this interpretation are the recent findings by Sanders et al. (38), who reported evidence suggesting that itch modulation via a single 30 sec period of skin cooling does not critically depend on the TRPM8 receptor. Although negative findings can occur for a variety of reasons, this may indicate that slight temperature changes are not be sufficient to trigger central modulatory itch systems through TRPM8. Sanders and colleagues suggest that other cold sensitive channels beyond TRPM8 could be involved in the itch-modulating effect of innocuous cold (39–41). Finally, it is also conceivable that the effect of transient skin cooling on histamine evoked itch is mediated by central inhibitory mechanisms. One possible mechanism of a top-down inhibition of itch could involve the descending inhibitory pathway originating in the periaqueductal gray (PAG), which is a well-known process in pain (for review, see 42). Carstens (43) found that histamine-induced discharge in dorsal horn neurons was strongly suppressed when the PAG was electrically stimulated, suggesting it may also play a role in top-down itch inhibition. Consistent with this idea, Mochizuki et al. (44) observed that activity in the PAG was increased when histamine evoked itch was suppressed by cold pain stimuli. However, more studies are needed to determine whether such top-down modulations can also account for thermal inhibition effects in the non-noxious range.
Regarding the effect of warming on histaminergic itch when focusing on comparisons within this condition at different time points, there is a tendency towards an anti-pruritic effect of warming, however, this was not reliable across participants. Inspection of individual time-courses indicated that 24% of participants (n = 4) reported a decrease of histaminergic itch upon skin warming, whereas others (n = 13) showed no systematic effect. Fruhstorfer et al. (1) observed a similar pattern, with 68% of participants showing an antipruritic effect of warming, but the remainder reporting the opposite (20%) or no systematic effect (12%). The effect of transient skin warming on histamine evoked itch could involve the Transient receptor potential vanilloid 4 (TRPV4) receptor, which responds to innocuous warming. In mice, it has been demonstrated that TRPV4 is expressed in subsets of sensory neurons that response to histamine (45). Pharmacological blockade of TRPV4 leads to reduced scratching to histamine, suggesting that TRPV4 is required for histamine-induced itch (45). In a very recent study by Sanders et al. (38), it was observed that a single period of a 30 s temperature change (either cooling to 18°C or warming to 38°C) decreased the scratching response to histamine evoked itch in mice. The effect of warming was blocked by a TRPV4 antagonist, suggesting that this receptor is causally involved in mediating the effect of warming on histamine evoked itch.
An interesting finding of the present study is the differential effect of transient skin warming on histamine vs. cowhage-induced itch. Whereas repeatedly warming the skin temporarily reduces histamine-induced itch (at least in a subset of participants), the same transient skin warming reliably enhances cowhage-induced itch. Since numerous physical factors could modulate terminal excitability or augment the receptor-agonist interaction, we can only speculate about the underlying mechanism at this point. One possible explanation is that since the activation of PAR2/4 receptors by cowhage is an enzymatic reaction (13), this process might be particularly affected by heat, as opposed to the activation of histamine receptors, which are G coupled Protein receptor types (46). Another aspect to consider is that the warm responses for the polymodal pruriceptive C-fibres conveying cowhage itch start at much lower temperatures than the mechanoinsensitive C-fibres (CMi) transmitting histamine induced itch (47–51). Thus, for histamine-induced itch, warmth might be providing counter-stimulation leading to a reduction in itch (at least in some participants), whereas for cowhage the same fibres are slightly more excitable during skin warming, resulting in an enhanced itch response (28, 52, 53).
In conclusion, the present study replicated previous findings of an itch-enhancing effect of short term skin cooling for histaminergic itch, and provided new evidence for an itch-enhancing effect of skin warming on cowhage-induced itch. More research is needed to better understand the neural mechanism underpinning such temperature-induced modulation of itch, both for histaminergic as well as non-histaminergic pathways. One way to gain further insights into the underlying mechanisms could be to test whether the effects of transient temperature change are strictly homotopic, or can also be elicited when thermal stimuli are applied distal to the skin region where itch was evoked (3).
This work was supported by a grant from the British Skin Foundation to HH and FC (project number: 7011s). We are grateful to Dr Ethan Lerner (Harvard Medical School) for kindly providing the cowhage spicules and Prof Gil Yosipovitch (University of Miami) for helpful comments on an earlier version of this manuscript.
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize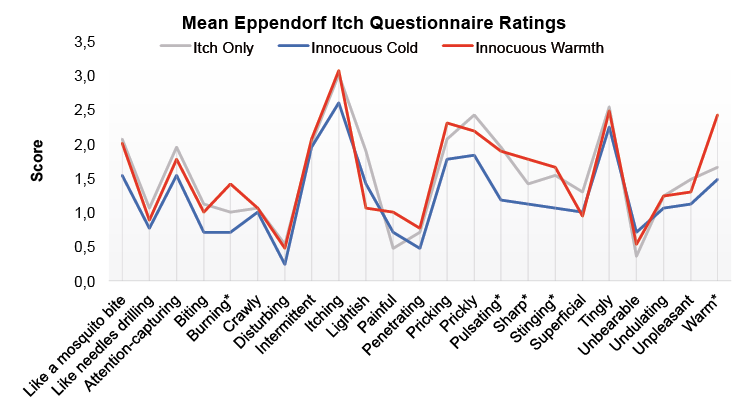 Click to show fullsize
Click to show fullsize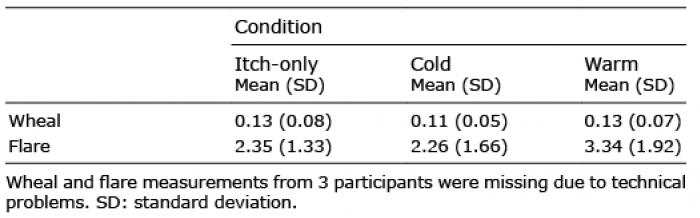 Click to show fullsize
Click to show fullsize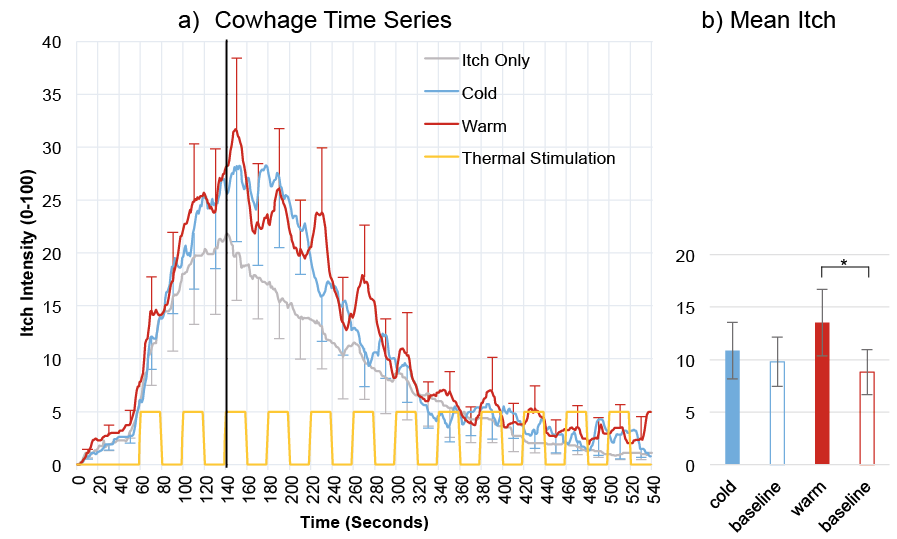 Click to show fullsize
Click to show fullsize