


Department of Dermatology, Aarhus University Hospital, Palle Juul-Jensens Boulevard 99, Aarhus N DK-8200, Denmark. *E-mail: Claus.Johansen@clin.au.dk
Accepted Aug 6, 2019; E-published Aug 6, 2019
Psoriasis is a common chronic inflammatory skin disease (1). Interleukin (IL)-17A has been identified as a key cytokine mediating the immunopathogenesis of psoriasis (2–4). However, the underlying molecular mechanism by which IL-17A mediates its psoriatic effects is unknown. A distinc-tive characteristic of psoriasis is the presence of neutrophils in the epidermis. Neutrophils are recruited to the epidermis by neutrophil-recruiting chemokines, such as C-X-C motif chemokine ligand (CXCL)1, CXCL5 and IL-8 produced by activated keratinocytes (5, 6). I kappa B kinase (IKK) ε is a protein encoded by the IKBKE gene. IKKε has been demonstrated to play an important role in the recruitment of neutrophils in IL-17A-induced inflammation (7). By the use of IKKε-deficient mice, it has been demonstrated that IKKε is required for IL-17A-induced lung inflammation and, interestingly, IKKε was found to play an important role in the recruitment of neutrophils (7). To date, the role of IKKε in psoriasis remains unknown; however, the above-mentioned data suggest that IKKε may play a role in the pathogenesis of psoriasis. The aim of the current study was to investigate the role of IKKε in psoriasis.
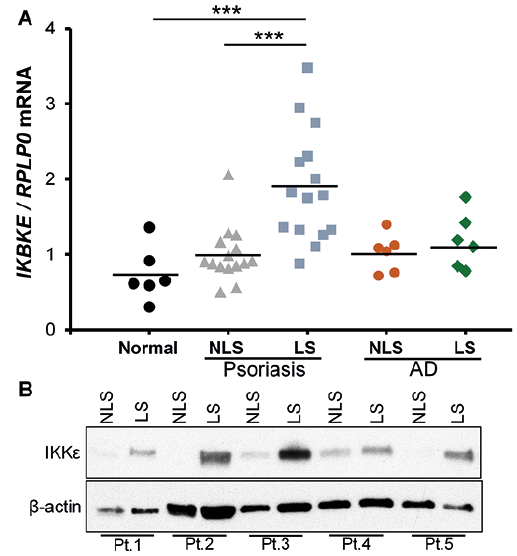
In order to examine IKBKE mRNA expression, RNA was isolated from punch biopsies obtained from both lesional and non-lesional skin from 15 psoriatic patients and 6 patients with atopic dermatitis. Furthermore, RNA was isolated from 6 healthy controls. The mRNA expression of IKBKE was significantly increased in lesional psoriatic skin compared with non-lesional psoriatic skin from the same patient; an approximately 2-fold increase was observed. The expression of IKBKE in lesional psoriatic skin was also significantly higher compared with healthy controls (Fig. 1A). To determine whether the observed increase in IKBKE expression was specific to psoriasis or simply due to increased inflammation in the skin we also investigated IKBKE expression in atopic dermatitis. No changes were found in IKBKE expression between lesional and non-lesional atopic dermatitis skin (Fig. 1A). It was also found that the IKKε protein level was increased in lesional psoriatic skin compared with non-lesional psoriatic skin from the same patient (Fig. 1B), supporting a potential role of IKKε in the pathogenesis of psoriasis.
Fig. 1. Increased levels of I kappa B kinase ε (IKKε) in lesional psoriatic skin. (A) RNA was extracted from lesional (LS) and non-lesional (NLS) skin biopsies from 15 patients with psoriasis, 6 patients with atopic dermatitis (AD) and from 6 healthy volunteers (Normal). IKBKE mRNA expression was analysed by quantitative PCR (qPCR). RPLP0 mRNA was used for normalization. The relative fold induction is presented as scatter plots. Data were analysed with 1-way analysis of variance (ANOVA). ***p < 0.001. (B) IKKε protein expression was examined in paired lesional (LS) and non-lesional (NLS) biopsies from 5 psoriatic patients (Pt.1–5) by Western blotting.
To characterize the role of IKKε in the regulation of specific psoriasis-associated genes, siRNA was used to knockdown IKKε. Transfection of human keratinocytes with IKKε siRNA significantly reduced the mRNA and protein expression of IKBKE/IKKε in tumour necrosis factor alpha (TNFα)- and/or IL-17A-stimulated cells compared with control siRNA transfected cells (Fig. S1). Notably, siRNA-mediated knockdown of IKKε, led to a minor, but significant, reduction in CCL20 mRNA expression after stimulation with IL-17A alone and with IL-17A in combination with TNFα, compared with control siRNA transfected cells (Fig. S2). Likewise, mRNA expression of DEFB4 and CXCL1 was significantly decreased in cells transfected with IKBKE siRNA, after combined stimulation with IL-17A and TNFα, but not when single stimulations were used (Fig. S2).
To further characterize the potential role of IKKε in the pathogenesis of psoriasis the imiquimod-induced psoriasis-like skin inflammation model was used. First, Ikbke mRNA expression in mice ears treated with imiquimod for 1, 3 and 5 days was investigated. Expression of Ikbke mRNA was increased significantly at both days 3 and 5 in imiquimod-treated mice compared with vehicle-treated mice (Fig. S3). To assess the functional role of IKKε in the imiquimod model, the model was applied on IKKε-deficient mice. IKKε deficiency in the mice was confirmed both by quantitative PCR (qPCR) and Western blotting (Fig. S4). Clinically, imiquimod-treated mice developed erythema and scaling of the skin. However, no differences were observed in the degree of erythema and scaling between IKKε-deficient and wild-type mice treated with imiquimod (Fig. S5). Imiquimod treatment caused weight loss of approximately 15% and increased the ear thickness by approximately 130%; however, no differences in ear thickness were observed between imiquimod-treated wild-type and IKKε-deficient mice (Fig. S6A, B). Haematoxylin and eosin (H&E) staining of skin biopsies revealed no histopathological differences between the 2 mice strains after treatment (Fig. S6C). Immunofluorescence staining revealed presence of neutrophils in skin sections after imiquimod treatment, however, no differences in the number of neutrophils between wild-type and IKKε-deficient mice were observed (Fig. S6D). Finally, the expression of psoriasis-associated genes was examined in the ear biopsies. Although imiquimod treatment increased Nfkbiz, Ccl20 and Cxcl1 mRNA expression compared with vehicle-treated mice, no alterations in Ccl20 and Cxcl1 mRNA expression were observed between imiquimod-treated wild-type and IKKε-deficient mice (Fig. S6E).
This study demonstrated an increased mRNA and protein expression of IKKε in lesional psoriatic skin. The increase in IKKε seemed to be specific for psoriasis and not just due to increased inflammation in the skin in general, because IKKε expression was unaltered between lesional and non-lesional skin from patients with atopic dermatitis. Previous data have shown that the function of IKKε in IL-17A-mediated signalling is linked with neutrophilia (7). IKKε was demonstrated to play a role in IL-17A-induced airway inflammation by regulating the expression of neutrophil-mobilizing cytokines and chemokines, including CXCL1 and CXCL2 (7). Here, we also found IKKε to be involved in the expression of CXCL1. However, only an involvement of IKKε on the CXCL1 expression was observed when TNFα and IL-17A were given in combination, whereas no involvement was found by stimulation with IL-17A alone. The discrepancy between our data and that of Bulek et al. (7) could be because the role of IKKε in IL-17A stimulation is cell-type specific.
To analyse how IKKε was regulated in vivo during psoriasis development, we used the imiquimod-induced psoriasis-like skin inflammation model, a well-described psoriasis mouse model (8). By comparing vehicle- and imiquimod-treated wild-type mice, we observed no increase in Ikbke mRNA over time during imiquimod treatment. Likewise, we demonstrated only a minor, although significant, induction of Ikbke mRNA upon imiquimod treatment at days 3 and 5. This is in contrast to what we observed in biopsies from psoriatic patients, where we demonstrated an almost 2-fold induction of IKBKE mRNA between lesional and non-lesional psoriatic skin. This could suggest that, in this specific psoriasis mouse model, or perhaps in mice in general, IKKε does not play a major role in the induction of psoriasis-like skin inflammation. Although the imiquimod-induced psoriasis mouse model is dependent on IL-17A signalling (8, 9), it is possible that other cytokines might play a more prominent role in mice, and therefore mice might not represent the best model to study IL-17A downstream effects. This theory was also supported by the fact that we observed no difference between Ikbke knockout mice and wild-type mice in their clinical appearance, bodyweight, ear thickness and expression of the inflammatory genes Ccl20 and Cxcl1 after imiquimod treatment. Surprisingly, IKKε deficiency significantly increased the imiquimod-induced Nfkbiz gene expression, suggesting that IKKε might have a protective role in this model.
Although we demonstrated increased expression of Ikbke mRNA in lesional psoriatic skin and that IKKε knockdown in vitro decreased IL-17A/TNFα-induced proinflammatory gene expression, this proinflammatory role of IKKε in psoriasis was not supported by results from the imiquimod-induced psoriasis mouse model. Thus, further studies are needed in order to fully elucidate the role of IKKε in psoriasis.
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize