


Departments of 1Pharmacology and Clinical Neuroscience, 3Odontology, Umeå University, and 2Clinical Chemistry, Region Va?sterbotten, SE-901 85 Umea?, Sweden. *E-mail: christopher.fowler@umu.se
Accepted Aug 8, 2019; E-published Aug 9, 2019
Oral lichen planus (OLP) is a chronic, inflammatory mucocutaneous disease characterized by the presence of subepithelial cytotoxic T cells. The resulting epithelial apoptosis causes great discomfort to the patient, and symptoms include burning sensations and sensitivity to spicy foods (1). Heat and spicy foods containing capsaicin produce their effects on sensory nerves by activation of transient potential receptor (vanilloid) 1 (TRPV1), and increased expression of TRPV1 receptors in the oral mucosa has been reported in OLP (2).
Oxylipins are a family of biologically active lipids produced by the oxidation of polyunsaturated fatty acids (3). The most well-known oxylipins are the prostaglandins, produced by cyclooxygenase (COX)-catalysed oxidation of arachidonic acid, and COX-2 expression is increased in OLP tissue compared with healthy biopsy tissue (see, for example, 4, 5). The role of linoleic acid-derived oxylipins is less well studied (and not at all in OLP), but there is evidence that they are involved in the pathogenesis of inflammatory pain, possibly through mechanism(s) involving activation of TRPV1 receptors (6, 7). Here we investigate linoleic acid-derived oxylipins present in biopsies from patients with OLP and controls, and report that the relative proportion of 9-hydroxyoctadecadienoic acid (9-HODE) to 13-HODE is increased 4-fold in OLP.
Biopsy samples, embedded in Tissue-Tek® and snap frozen, were used. Seven linoleic acid derivatives were identified in the samples (Fig. 1). Note that the method used did not involve chiral separation of the lipids into R- and S-forms. The embedding material did not contain detectable levels of these lipids.
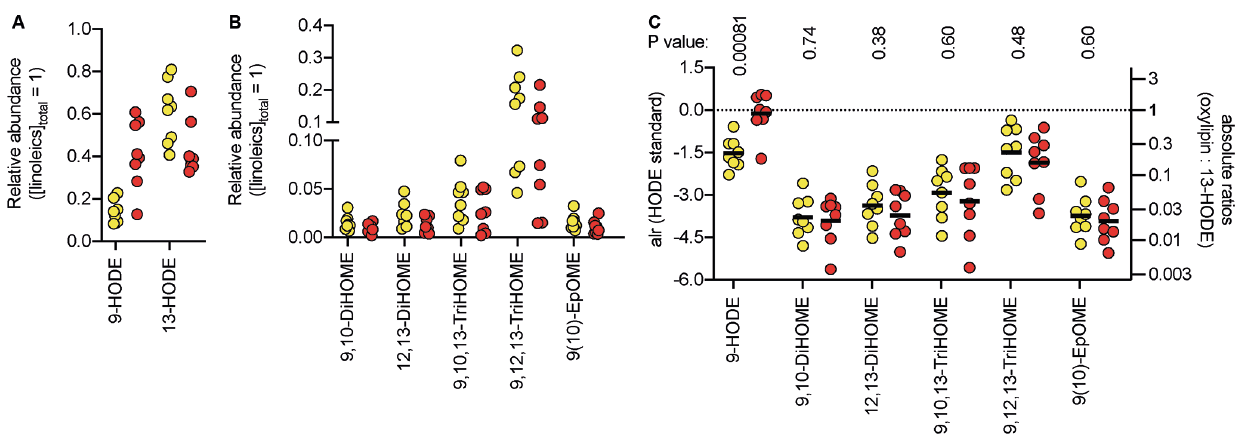
Because the sample sizes in the embedded materials were not known, absolute levels per unit wet weight could not be calculated. However, the proportions of each lipid to the total amount of the linoleic acid derivatives detected could be calculated for each sample, where it was seen that 9- and 13-HODE are predominant (Fig. 1A, B). Statistical evaluation of this type of data, termed compositional data, uses log-ratio transformations (8). The simplest of these, the additive log-ratio (alr, 8) has been used here using the most abundant lipid (13-HODE) as the reference (Fig. 1C). A multivariate analysis of variance (MANOVA) of the data gave a significant effect of patient group (using Pillai’s trace, V = 0.74, F6,9 = 4.25, p = 0.026). The log-ratio of 9-HODE/13-HODE was significantly greater in OLP than in the controls, whereas the other 5 ratios were not significantly different from the controls. Using the geometric means for the anti-logged values, the ratio of 9-:13-HODE was 4.1-fold (95% confidence intervall 2.0–8.2) higher in the OLP biopsies than in the control biopsies. An alternate approach, using alr values with 9-HODE as the reference, gave significant p-values for all the lipid ratios (Fig. S1). Thus, the data-set shows a changed concentration of 9-HODE relative to the other 6 linoleic acid derivatives.
Fig. 1. Proportions of linoleic acid-derived oxylipins in biopsies from 8 healthy controls (yellow) and from 8 oral lichen planus (OLP) patients (red). (A and B) Fractional contents, i.e. the concentration of lipid/combined concentration of all 7 lipids. Note the different scales on the y-axis in the panels. (C) Data are expressed as the additive log ratio (alr) with 13-hydroxy-9Z,11E-octadecadienoic acid (13-HODE) as the denominator, i.e. ln(lipid/[13-HODE]), with the anti-logged absolute ratios shown on the right y-axis. In all cases, the alr distributions passed the Shapiro–Wilk normality test, and the solid bars represent the means. The p-values refer to Welch t-tests (i.e. not assuming equal variances). The p-values are all unadjusted for multiple testing, but the critical value of p assuming a 5% false discovery rate was 0.0083. Although residual plots were deemed to be acceptable, it can be argued that non-parametric tests are more appropriate when sample sizes are small. For the 9-: 13-HODE log ratio, a Mann–Whitney U test gave a p-value of 0.0030. The corresponding p-values for the other log ratios were all non-significant. Lipid abbreviations: see Appendix S1.
This study found a large change in the relative proportions of 9-HODE with respect to 13-HODE in biopsies obtained from patients with OLP. Two questions arise: (i) by which enzymic pathway is this increased proportion of 9-HODE relative to 13-HODE seen; and (ii) by which mechanisms could such a change contribute to the pathogenesis of OLP.
With respect to (i) an obvious candidate is COX-2, since this enzyme converts linoleic acid to hydroperoxy-linoleates (HPODEs), which are then reduced to HODEs (9). Increased COX-2 activity is a hallmark of OLP (4, 5). However, both 9- and 13-HODE are produced by this pathway (9), and so a change in the relative proportions of 9-HODE and 13-HODE would not be expected. An alternative candidate enzyme is 12R-lipoxygenase, found in skin and tonsils (10, 11). Altered expression or mutations of this enzyme are associated with skin disorders (11). Given that ALOX12B, which codes for 12R-lipoxygenase, is upregulated in the epithelium of biopsy samples from patients with OLP (12), the changed log ratio of 9-HODE: 13-HODE found here may reflect a changed 12R-lipoxygenase activity.
With respect to (ii), it must be borne in mind that a changed ratio of 9-HODE:13-HODE does not necessarily mean that 9-HODE levels are higher, since the same result would be found if 13-HODE levels were decreased, or a combination of both of these changes was in play. If it is assumed, for the sake of argument, that the results reflect increased 9-HODE levels, then several possible downstream effects can be considered. Both enantiomers of 9-HODE have been reported to disrupt epidermal barriers in human epidermal keratinocyte cultures (13), although the concentration used (10 µM) was high. One of the targets of 9-HODE is the orphan receptor G2A (GPR132), and mice deficient in this enzyme show reduced mechanical pain thresholds following repeated treatment with oxiliplatin (14). Interestingly, incubation of dorsal root ganglion neurones with sub-micromolar concentrations of 9-HODE sensitize the response to the TRPV1 agonist capsaicin (14). GPR132 has a wide expression, including in skin (15). Re-investigation of the raw data from the genome-wide investigation of biopsy samples (12) indicated that GPR132 expression levels in the OLP samples are not significantly different from healthy controls (Table SI). Thus, the target for 9-HODE is present in OLP, at least at the level of mRNA. It is thus possible that the burning and painful symptoms reported by patients with OLP may involve 9-HODE-induced sensitization of TRPV1 receptors.
In conclusion, we report here, for the first time, that the balance of 9-HODE:13-HODE is changed in biopsy samples from OLP. The result may reflect a change in the expression of 12R-lipoxygenase and may contribute to the symptomatology of the disorder, the operative word here being “may”. Further studies using chiral separation of the HODE enantiomers and determination of their absolute, as opposed to relative, levels, as well as immunohistochemical and/or activity measurements of 12R-lipoxygenase in OLP, would shed light on these potentially important possibilities.
The authors thank Region Västerbotten and the research funds of the Medical faculty of Umeå University for their financial support of our work into OLP.


 Click to show fullsize
Click to show fullsize