


1Department of Molecular Biomedical Sciences, 2Comparative Medicine Institute, and 3Department of Clinical Sciences, North Carolina State University, Raleigh, NC, USA
Most canine visits to veterinarians are related to skin diseases with itch being the chief complaint. Historically, several itch-inducing molecules and pathways have been identified in mice, but whether or not these are similar in dogs is not yet known. Herein, we set out to study the expression of pruritogenic neuropeptides, their cognate receptors with a limited functional validation thereof using a multidisciplinary approach. We demonstrated the expression of somatostatin and other major neuropeptides and receptors in canine dorsal root ganglia neurons. Next, we showed that interleukin-31, serotonin, and histamine activate such neurons. Furthermore, we demonstrated the physiological release of somatostatin from dog dorsal root ganglia neurons in response to several endogenous itch mediators. In summary, our results provide the first evidence that dogs use similar pruritogenic pathways to those characterized in mice and we thus identify multiple targets for the future treatment of itch in dogs.
Key words: dog; canine atopic dermatitis; somatostatin; NPPB; NMB; GRP.
Accepted Aug 22, 2019; E-published Aug 22, 2019
Acta Derm Venereol
Corr: Santosh K. Mishra, Department of Molecular Biomedical Sciences, North Carolina State University, 1060 William Moore Drive, Office 242, Raleigh, NC 27607, USA. E-mail: skmishra@ncsu.edu
In the last few years, molecular studies have characterized multiple itch mediators, receptors, and neurotransmitters/neuropeptides that are required for itch signaling in mice. However, it is unclear if similar signaling components and pathways are involved in the propagation of itch in dogs. Herein, we focus on the physiological expression and function of itch-associated receptors and neurotransmitters that were first implicated in generating the itch sensation in mice. Our work identifies several new potential targets for antipruritic therapies in dogs.
Outside of preventative health care, dermatological issues account for nearly 20% of all causes for dog veterinary visits; approximately 30% of which are due to pruritus, canine atopic dermatitis being one of the leading causes of such pruritus (1). Research aimed at understanding the etiology of pruritus in dogs has begun with the characterization of receptors for several common pruritic agents found to be expressed in dog dorsal root ganglia (DRG) (2). In mice, a few neuropeptides are released from the DRG into the spinal cord synapses upon a cutaneous pruritic challenge: the natriuretic precursor peptide B (NPPB) (3), somatostatin (SST) (4), neuromedin B (NMB) (5) and the gastrin-releasing peptide (GRP) (6). However, itch-associated neurotransmitters/neuropeptides and their respective receptors have not yet been reported in dogs.
Here, we used qRT-PCR to determine the mRNA levels of itch-associated neuropeptides and neuropeptide receptors expressed in dog DRGs and spinal cords. Additionally, we used immunohistochemistry (IHC) to localize the expression of the SST protein in DRGs. Next, we performed calcium imaging on primary cultured dog DRG neurons to identify if they have functional receptors for selected itch mediators. Finally, we performed enzyme-linked immunosorbent assay (ELISA) on the supernatants from primary cultured dog DRG neurons stimulated with itch mediators to demonstrate the subsequent release of the itch-associated neuropeptide SST.
RNA isolation
RNA was isolated from fresh-frozen lumbar L4 and L5 DRG from 3 to 5 euthanized Beagles (3 females, 1-year-old; and 2 males, 7-year-old, 27 ± 2.0 kg, all from Marshall Farms). DRG had been flash frozen or placed in RNA later and stored at –80°C until use. RNA was similarly isolated from the lumbar spinal cords from 3 adult beagles. Dogs had no prior history of any known pain or itch conditions. Both DRG and spinal cord tissues were isolated under protocols approved by NC State’s animal care and use policies for different studies. A roughly 2 mm3 piece was cut from each DRG and transferred to a 10 ml tube containing 3 ml lysis buffer with 1% β-mercaptoethanol from Qiagen’s RNeasy Fibrous Tissue Mini Kit and then homogenized for 30–45 s. For the spinal cord, a length of ~4 mm was transferred to a 10 ml tube containing 3 ml of lysis butter with 1% β-mercaptoethanol from Qiagen’s RNeasy Fibrous Tissue Mini Kit and then homogenized for 30–45 s. Following homogenization, RNA was extracted following Qiagen’s protocol without performing a proteinase K digestion.
cDNA synthesis
To synthesize cDNA 200 ng RNA (in ≤ 8 µl), 2 µl random hexamer primers (Invitrogen, Waltham, MA, USA) and PCR grade water (for a final volume of 10 µl) was heated at 72°C for 3 min. Afterwards, 4 µl of 5x Reaction Buffer, 2 µl DTT, 2 µl SmartScribe Reverse Transcriptase (Clontech, Mountain View, CA, USA), and 2 µl of a 10 mM dNTP stock (Roche, Branchburg, NJ, USA) were added and the mixture was incubated at 42°C for 90 min followed by 72°C for 15 min and stored at –20°C until needed.
Quantitative real-time PCR
The following Taqman probes for the genes studied in this work were purchased from Thermofisher, Carlsbad, CA, USA: NPPA (Cf04393960), NPPB (Cf04393958), NPPC (Cf02714191), NPR1 (Cf02655890), NMB (Cf02713174), NMBR (Cf02697155), SST (Cf02995138), SSTR1 (Cf02695139), SSTR2 (Cf02695151) and GAPDH (Cf04419463). Custom Taqman probes were generated for GRP and GRPR using Thermofisher’s custom design software with bioinformatic analysis. The NCBI deposited sequences used to generate the custom Taqman Probes were: XM_861026.5 (GRP) and XM_014111958.2 (GRPR). GRP had two possible Taqman probe sets and we selected the 96 bp amplicon Taqman set. All samples were run on an Applied Biosystems StepOnePlus Real Time PCR System using Taqman Gene Expression Master Mix (Applied Biosystems, Waltham, MA, USA, 4369016) with the recommended qPCR cycle. CT values were calculated using StepOne Software v2.2.2 (Applied Biosystems). GAPDH was used as a housekeeping gene for normalization. A minimum of two technical replicates were used for each gene in the DRG and 3 technical replicates were used for each gene in the spinal cord. Samples from 5 dogs were used to test for NPPA, NPPB, NPPC, and NPR1; from 3 dogs were used to test for NMBR, SSTR1, and SSTR2; and 4 dogs were used to test for GRP and GRPR in the DRG. All genes were tested in 3 dogs in the spinal cord. Relative tissue expression values were calculated using the following equation: relative expression = 2-ΔCT, where ?Ct =Ct, GAPDH−Ct, GOI; where GOI is the gene of interest. The graphs depict the median values. The median for each gene is shown in the results section.
Immunohistochemistry
Pieces of fresh frozen DRGs were placed in Tissue-Tek (Sakura, 4583) and sectioned to 10–14 µm thickness. Sections were placed onto Superfrost Plus Gold Microscope slides (Fisher, 15-188-48). Following sectioning, slides were placed in a 4% PFA (Sigma, St. Louis, MO, USA, P6148) for 10 min, washed with 1X PBS 3 times, and then stored at room temperature for 60 min with a 5% BSA (VWR, 0332-100G; w/v), 0.1% Triton-X100 (Sigma, T8787; v/v) blocking solution. The slides were then washed in 1X PBS 3 more times and then stored overnight at 4°C with the primary antibody in a 5% BSA, 0.1% Triton-X100 solution. The following day, slides were washed 3 times in 1X PBS and then incubated at room temperature for 60 min in a 2% BSA, 0.01% Triton-X100 solution with the secondary antibody. Slides were then washed 3 times in 1X PBS, vacuum dried, and mounted with Prolong Diamond Antifade mountant with DAPI (Thermofisher, P36962). Slides were stored at 4°C in the dark until imaging on a Nikon TE200 microscope. The following primary antibody was used to stain for SST: rat anti-human SST (abcam, ab30788; 1:500, which was selected because human and dog SST are 98% identical according to an alignment run using NCBI’s BLASTp program. The following secondary antibody was used at a 1:200 dilution goat anti-rat IgG conjugated to Cy3 (Life Technologies, Waltham, MA, USA, A10522).
Dorsal root ganglia collection and dissociation for enzyme-linked immunosorbent assay and calcium imaging
DRG from euthanized shelter dogs (two pit bull crossbred dogs, one Labrador crossbred, and one basenji crossbred, all adult spayed females of unknown age) were collected within 5 h of euthanasia. During isolation, DRG were kept in ice cold HBSS without calcium or magnesium (VWR, Suwanne, GA, USA, 02-0121). DRG were then moved to dissociation media containing 5 mg/ml collagenase type 3 (Fisher, Waltham, MA, USA, 150704) and 5 U/ml dispase (VWR, 42613-33-2) and incubated for 60 min at 37°C with 5% CO2. The tissue was then disrupted using an autoclaved 1 mL pipette tip with the bottom ~0.5 cm cut off attached to a pipette set to 1 mL. The DRG were then placed back in the incubator for 45 min at 37°C with 5% CO2. The DRG were disrupted again with an autoclaved 1 ml pipette tip with the bottom ~0.2 mm cut off attached to a pipette set to 1 ml. Following this, the DRG were split into two 1.5 ml Eppendorf tubes and placed into a 37°C water bath for 30 min so that the debris could settle into the bottom of the tube and any fat at the top of the tube. After this incubation, the middle layer containing cells was moved into a 50 ml conical tube containing 5 ml of 12% BSA (VWR, 0332-100G) and then centrifuged for 15 min at 1,000 rpm. The supernatant was removed, and the cells were resuspended in 10 ml of complete media (DMEM with 10% FBS (VWR, 97068-085) with 1% PenStrep (Millipore, Burlington, MA, USA, TMS-AB2-C) and then centrifuged for 3 min at 1,000 rpm. The supernatant was removed and then the cells were resuspended in 10 ml of fresh complete media, spun down for 3 min at 1,000 rpm. The previous step was repeated one more time for a total of 3 resuspensions and spin downs.
Enzyme-linked immunosorbent assay (ELISA)
After dissociation, cells were resuspended in 1 ml complete media and counted on a hemocytometer. For each compound tested approximately 10,000 cells/dish were added to wells that had been coated with a solution containing poly-L-lysine (Sigma, P4707) and 100 µg/ml laminin (Sigma, L2020) in a 24-well plate. Two dishes/per dog was tested for each mediators. Complete media was then added so that the final volume in each well was 1 ml. Cells were then incubated overnight at 37°C with 5% CO2. Cells were stimulated with either 0.3 μM canine IL-31 (Novus Biologicals, Centennial, CO, USA, NBP2-59591), 100 μM serotonin (Sigma, H9523), 50 μM histamine (Sigma, H7125), or vehicle (DMEM) with and without cells. Cells were incubated for 8 h at 37°C with 5% CO2. This time point was selected because it has produced the maximal response in other ELISA assays using 0.3 μM IL-31 (7, 8). The serotonin and histamine concentrations are based on the concentration used for calcium imaging (9, 10). After this incubation, the supernatant was removed and stored immediately at –80°C. The amount of SST released from these cells was measured using the ELISA protocol for LSBio’s Dog SST ELISA kit (LS-F14876).
Calcium imaging
The remaining cells were plated on coverslips coated with the same laminin and poly-L-lysine coating. Cells were incubated for 3 h at 37°C with 5% CO2. Following this incubation 1 ml of complete media was added and then the cells were incubated overnight at 37°C with 5% CO2. Before imaging, cells were incubated with 4.2 µM Fura-2AM (Enzo, Farmingdale, NY, USA, ENZ-52006) for 30 min at 37°C with 5% CO2. During imaging, the cells were perfused with Locke buffer containing the following: 135 mM sodium chloride, 3.2 mM potassium chloride, 2.5 mM magnesium chloride, 2.8 mM calcium chloride, 667 µM monobasic sodium phosphate, 14.2 mM sodium bicarbonate, and 10.9 mM D-glucose (all VWR) with a pH between 7.00 and 7.40. The buffer and the holding plate were kept at 37°C while imaging. All compounds were added directly to the perfusion buffer in the following concentrations: 0.3 µM IL-31, 100 µM serotonin, 50 µM histamine, 1 µM capsaicin, and 1 mM KCl. Imaging data was collected on a TE200 inverted microscope using NIS Elements software (Nikon). Cells were exposed to 340 nm and 380 nm wavelengths for 300 ms and the A340/A380 ratio was calculated. Cells not responding to KCl, which was applied at the end of each experiment, were regarded as non-neuronal cells and excluded from analysis. Traces were analyzed using Excel and responses greater than 10% of the baseline were counted. Error bars represent ± SD.
Statistical tests
Statistical tests were run using Prism (Graphpad) version 8.0.2. Because there were fewer than 10 samples from dogs with mixed-breed backgrounds, a Shapiro-Wilk test was run to determine if the calcium imaging and ELISA data followed a normal distribution. This test determined that the distributions of the calcium imaging data results for IL-31, serotonin, histamine, and capsaicin were normally distributed. Further, Grubb’s test for outliers with α = 0.05 found that none of the results was an outlier. Therefore, all calcium imaging data points were included. The Shapiro-Wilk test determined that the distribution of the ELISA results for vehicle, IL-31, serotonin, and histamine were not normally distributed. Therefore, the ELISA data was analyzed using a one-tailed Mann-Whitney U test with individual comparisons between the vehicle-treated cells and cells treated with IL-31, serotonin, and histamine.
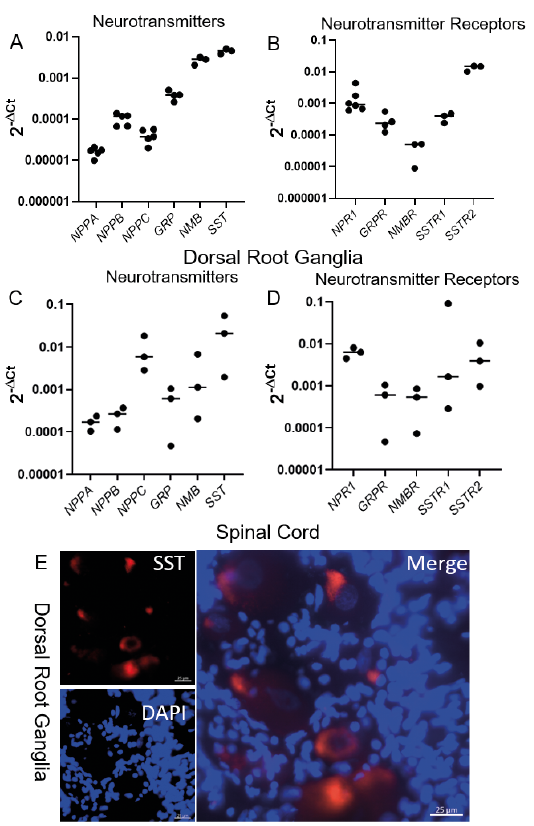
We found that dog DRGs produced mRNA coding for the itch-associated neuropeptides NPPB (0.67 to 1.2 × 10–4), GRP (2.6 to 5.1 × 10–4), NMB (2.05 to 3.18 × 10–3), and SST (3.84 to 5.11 × 10–3) (Fig. 1A). Additionally, we found a minimal expression of NPPA (0.99 to 2.09 × 10–5) and NPPC (2.0 to 5.7 × 10–5), two neuropeptides in the natriuretic family. We also discovered that dog DRGs expressed mRNA coding for NPR1 (0.60 to 4.43 × 10–3), GRPR (1.2 to 5.5 × 10–4), SSTR1 (2.4 to 4.0 × 10–4) and SSTR2 (1.02 to 1.54 × 10–2) (Fig. 1B) with relatively small levels of expression of NMBR (0.88 to 5.2 × 10–5).
Since these neuropeptides are released centrally and activate receptors expressed on spinal interneurons/projection neurons, we also performed qRT-PCR on normal dog spinal cord samples. There, we detected a positive expression of the itch-associated neuropeptides SST (0.2 to 5.4 × 10–2) and NMB (0.2 to 6.7 × 10–3), with only a minimal mRNA expression of NPPA (1.04 to 2.38 × 10–4) and NPPB (1.15 to 3.68 × 10–4) (Fig. 1C). Interestingly, transcripts levels of NPPC (0.28 to 1.82 × 10–2) were similar to those of NPR1 in the spinal cord. Finally, we detected a positive expression of mRNA encoding for the neuropeptide receptors NPR1 (4.5 to 8.1 × 10–3), NMBR (0.7 to 8.6 × 10–4) and both SSTR1 (0.02 to 9.1 × 10–2) and SSTR2 (0.1 to 1.0 × 10–2) (Fig. 1D). As in mice (5, 6), the expression of mRNA for both GRP (0.05 to 1.0 × 10–3) and its receptor GRPR (0.04 to 1.0 × 10–3) was also found in the dog spinal cord (Fig. 1D). Since our qRT-PCR demonstrated that SST was the most abundantly expressed itch-associated neuropeptide at the mRNA level in the dog DRGs, we performed immunohistochemistry for this neuropeptide on sectioned dog DRGs: as expected, we detected SST-positive neurons therein (Fig. 1E).
Fig. 1. Expression of itch associated neurotransmitters and their receptors in canine dorsal root ganglia (DRG) and spinal cord. A) Relative mRNA expression of itch neurotransmitters in the DRG. B) Relative mRNA expression of itch neurotransmitter receptors in the DRG. C) Relative mRNA expression of itch neurotransmitters in the spinal cord. D) Relative mRNA expression of itch neurotransmitter receptors in the spinal cord. Results for qPCR are displayed as individual points with a bar denoting the median value. E) Representative image showing somatostatin positive neurons (red) with DAPI (blue). Biological replicates are represented by a dot on scatter plot which is a mean of two technical replicates (DRG) or 3 technical replicates (spinal cord) for all genes tested. The median value was highlighted by the bar in panels A–D.
Next, we studied if itch mediators induced a calcium influx in dog DRG neurons. Interestingly, we found that IL-31, serotonin, histamine, and capsaicin were all capable of causing calcium influx into these sensory neurons (representative trace; Fig. 2A). A quantitative study demonstrated that IL-31 induced a calcium influx in 7.7 ± 2.5% (12/163), serotonin in 5.6 ± 5.3% (9/163), histamine in 13.6 ± 4.2% (21/163) and capsaicin in 27.0 ± 6.0% (42/163) of such neurons, respectively (Fig. 2B). All 3 endogenous mediators IL-31, histamine, and serotonin were within the capsaicin-positive subset of canine DRG neurons (Fig. 2C).
Finally, we studied the physiological release of the itch-associated neuropeptide SST in response to itch mediators, as, in mice, both SST and NPPB are co-expressed almost exclusively with each other and we found a similar expression of SST in dog DRGs, We thus stimulated a primary culture of dog DRG neurons with IL-31, serotonin, and histamine (Fig. 2D). We found that stimulating dog DRG neurons with vehicle did not cause a significant increase in SST release (7.8 ± 6.8 pg/ml) when compared to no stimulation (0.6 ± 1.1 pg/ml). A challenge with IL-31, serotonin, and histamine caused a significant release in the amount of SST released (58.2 ± 32.5 pg/ml, p = 0.014 and 56.8 ± 38.2 pg/ml, p = 0.014; 64.0 ± 15.0 pg/ml, p = 0.014 respectively).
Fig. 2. Itch-inducing compounds are capable of activating primary cultured canine dorsal root ganglia (DRG). A) Representative trace of a neuron responding to interleukin (IL)-31, histamine, capsaicin and KCl. B) Quantification of neurons responding to IL-31 (0.3 µM), serotonin (100 µM), histamine (50 µM), capsaicin (1 µM), each dot in the scatter plot represents single dog. C) Venn diagram represents overlapping population, n = 4, nneuron = 39, 32, 60, 32 for a total of 163 neurons. D) Stimulating primary culture dog DRG neurons with IL-31 (0.3 µM), serotonin (100 µM), histamine (50 µM) causes a significant increase in the amount of somatostatin released as compared to vehicle (both with and without cells). Data is presented as mean ± SD. n = 4 dogs, ns: not significant, *p = 0.014 one-tailed Mann-Whitney U Test.
This appears to be the first study that reports the expression of common itch-associated neurotransmitters/neuropeptides and their receptors in the normal dog. Calcium imaging studies on primary culture neurons isolated from euthanized dogs demonstrated functional receptors for IL-31, serotonin, and histamine. Additionally, we showed that itch mediators induce neuropeptide release from dog DRG neurons in culture and we thus provided evidence of neurotransmitter-induced itch propagation.
We found a positive expression of NPPA, NPPB, NPPC, GRP, NMB, and SST in dog DRGs. Our data illustrated that NPPA and NPPC had the lowest levels of expression, followed by NPPB, GRP, NMB and SST having the highest. These relatively low expression levels of NPPB, GRP and NMB do not exclude the involvement of these neuropeptides in the neurotransmission of itch, as has been demonstrated in mice (3, 5, 6). However, as we found a minimal expression of NMBR in the spinal cord, it is unclear if the dogs utilize an NMB spinal itch pathway as in mice. The role of NPPA and NPPC in DRG-to-spinal cord signaling transmission is unknown in dogs as in mice.
Interestingly, we also detected NPR1, SSTR1 and SSTR2 mRNA in the normal dog DRG. SSTR2 has been shown to be expressed in human and mouse DRGs, both at the protein (11) and mRNA levels (12), and this opens the possibility of an autocrine amplification loop involving SST and SSTR2 within the DRG upon peripheral stimulation. An important limitation of our methodology, however, is that we extracted total RNA from whole dog DRGs, which means that we captured RNA from both neuronal but also non-neuronal populations. Hence, it is possible that some of the transcripts detected might have been from the DRG-associated satellite glial cells, fibroblasts, macrophages, and other cell types present in these ganglia. Based on these results, it is likely that dogs use these neurotransmitters in a yet-unknown combination to generate itch signals that are later propagated to the central nervous system.
In the dog spinal cord, we unexpectedly found a positive expression of mRNA encoding for NPPC at a transcription level similar to that of NPR1 in the spinal cord. This natriuretic peptide has been shown to modulate the activity of TRPV1 in the DRG (13). Further research is needed to determine the role of NPPC in the sensory transmission of itch and pain in the dog.
Our calcium imaging results demonstrated that the percentage of neurons responding to serotonin, histamine, and capsaicin was similar to that reported in mice (9, 10, 14, 15). Interestingly, we found that the percentage of neurons responding to IL-31 in dogs was roughly twice as large as that reported for mice (8) and humans. While dogs mount a strong pruritic response when challenged with IL-31 (16), they do not when provoked with histamine or serotonin (17). This difference is hard to explain as dog DRG neurons respond with a calcium influx when challenged with both histamine and serotonin as for IL-31. There are, in fact, more neurons responding to the formers than the latter. It is possible that dogs are similar to mice wherein the percentage of neurons responding to a pruritogen does not correlate with the behavior response (9). Future studies should address this dilemma. Either way, these results are compatible with earlier reports of dog DRG neurons expressing the IL31Rα (18).
We demonstrated the release of SST in primary culture dog DRGs. We found that IL-31, serotonin, and histamine treated cells were all capable of causing a significant increase in the amount of SST released as compared to vehicle treated cells.
In summary, our results demonstrate that the generation of itch sensation in dogs might utilize multiple neuronal pathways from DRGs to the spinal cord, similar to those found in mice. Our study opens the door for the development of new therapeutics by using combinatorial approaches for the treatment of canine pruritus associated with atopic dermatitis and other forms of intractable pruritus in dogs.
Funding for this project was provided by start-up funds awarded to SKM from NC State University. JJW’s scholarship support is a gift from Dr. John M. Davis, University of Chicago. The authors would like to thank Drs. Constanza Meneses, John Hash, and Joy Ganchingco for providing and helping with harvesting spinal cord and DRG tissue and for DRG dissociation protocol.
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize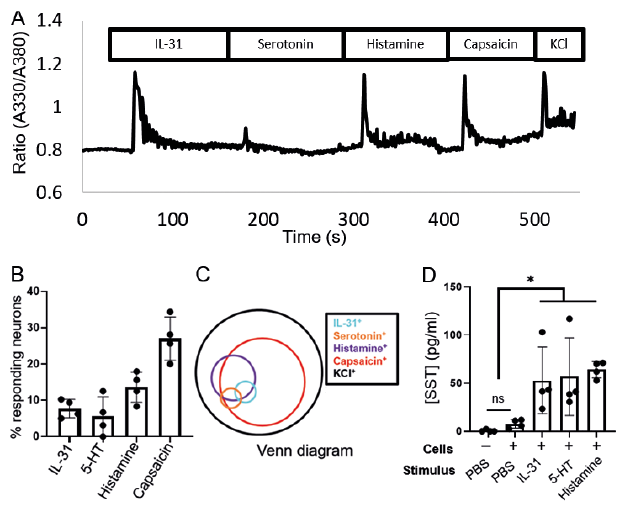 Click to show fullsize
Click to show fullsize