


Department of Dermatology, Shinshu University School of Medicine, 3-1-1 Asahi, Matsumoto 390-8621, Japan. *E-mail: rokuyama@shinshu-u.ac.jp
Accepted Dec 10, 2019; Epub ahead of print Dec 10, 2019
Acta Derm Venereol 2020; 100: adv00045
The advent of mutated BRAF-targeted therapy has revolutionized the treatment landscape of melanoma, but the clinical benefits of BRAF inhibitors are limited by intrinsic and acquired resistance. Tumour heterogeneity, the presence of both BRAF-mutated cells and BRAF-wild-type cells, may contribute to acquisition of resistance to targeted treatment (1, 2). This study aims to elucidate the heterogeneous nature of BRAF genotypes of primary tumours and metastatic lesions.
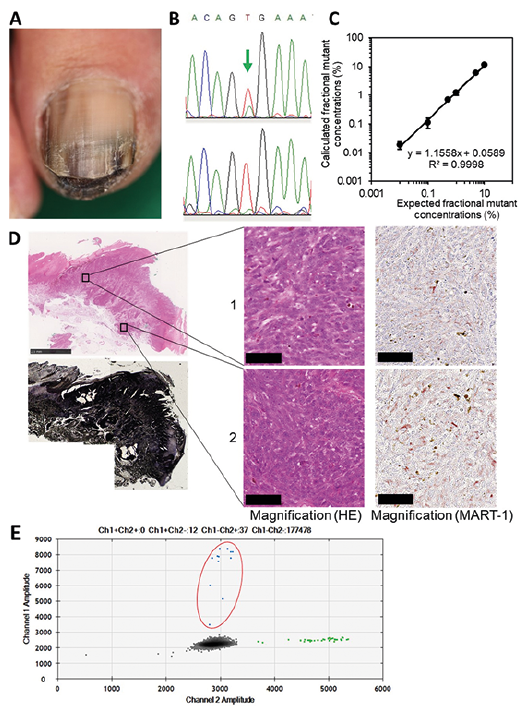
A 49-year-old Japanese woman presented with a dark-brown nodule on her toenail and inguinal lymph node metastases (Fig. 1A). Melanoma was diagnosed and she was treated by surgical resection. Shortly thereafter, metastases appeared in the lung, brain, bone and skin. Sanger sequencing revealed differences in BRAF genotypes: BRAFV600E in the primary tumour and BRAFwild in a skin metastatic lesion (Case 1, Table I). However, the BRAFV600E allele was rare in the primary tumour, as reflected by the small size of the mutation peak in the sequence chromatogram (Fig. 1B). Importantly, because Sanger sequencing does not detect genotypes present in less than 15–20% of cells (3), the BRAFV600E allele may have been present at trace levels in the skin metastatic lesion. Therefore, the BRAFV600E allele frequency (AF) was measured quantitatively by droplet digital PCR (ddPCR) (QX200 ddPCR system, Bio-Rad, Hercules, CA, USA) (4). The detection limit of ddPCR was 0.01% BRAFV600E per 100 ng of DNA; moreover, the assay was linear over the entire test range (Fig. 1C).
The presence of stromal and inflammatory cells can affect the AF in a sample. To minimize this potential confounder, we chose 2 sites containing many melanoma cells and few non-melanoma cells, and then dissected them using a laser-capture microdissection system (Fig. 1D). For the ddPCR, all specimens were run in duplicate. The mean BRAFV600E AF at the 2 sites was 17.6% and 19.3%, respectively (Fig. 1E and Table I). By contrast, MART-1 (a melanoma marker) positivity was 92.1% and 89.2%, respectively, as determined by immunohistochemistry. Contamination by BRAFwild derived from non-melanoma cells cannot explain the discrepancy between the low BRAFV600E AF and the high proportion of melanoma cells at the 2 sites. Furthermore, the metastatic skin lesion had a BRAFV600E AF of 0.02% (Table I). Thus, the BRAFV600E allele was very rare, but present, in the metastatic lesion, a finding consistent with the heterogeneity of melanoma.
Next, to analyse the heterogeneity of BRAF genotypes, we investigated whether there were cases in which the BRAFwild genotype was detected in metastatic lesions even when BRAFV600E was detected in primary tumours. Using Sanger sequencing, we analysed BRAF genotypes in metastatic lesions of another 8 cases with BRAFV600E-primary tumours (cases 2–9). Two of these (cases 3 and 8) were characterized as harbouring BRAFwild in metastatic lesions (Table I). Because Sanger sequencing is not very sensitive (3), ddPCR was used to analyse the BRAFV600E AF in primary and metastatic lesions from both cases. As in case 1, we then performed microdissection to isolate melanoma lesions containing few non-melanoma cells. In cases 3 and 8, MART-1 positivity was 82.9–88.9% and 75.8–90.1%, respectively. The BRAFV600E AF in primary tumours was high (case 3, 68.9%, 74.9%, 78.6% at 3 sites; case 8, 31.0%, 34.9%, 37.3% at 3 sites), but low in metastatic lesions: 1.28% in case 3 and 0.19% in case 8 (Table I). Thus, although the BRAFV600E allele was abundant in the primary tumours, it was rare in metastatic lesions. These results can be explained by BRAF heterogeneity, but not by contamination by non-melanoma cells.
Fig. 1. (A) Clinical features of the primary tumour. (B) Sequence chromatogram of DNA from the primary tumour (upper) and skin metastasis (lower). The arrow shows the BRAFV600E peak. (C) Sensitivity of the ddPCR assay of BRAFV600E. BRAFV600E DNA was diluted in BRAFwild DNA (total DNA=100 ng). To validate the limits of detection, BRAFwild DNA isolated from Mel-2 cells (BRAFwild homozygote) was spiked with BRAFV600E DNA isolated from A375 human melanoma cells (BRAFV600E homozygote) and then the percentage of BRAFV600E DNA measured. Assays were performed in triplicate. Spearman’s rank correlation coefficient: R2 = 0.9998, p < 0.01. (D) Histopathological and immunohistological images of the primary tumour in case 1. Upper left, haematoxylin/eosin stain; lower left, tissue after microdissection; right, microdissected tissues. Scale bars: 2.5 mm (upper left) and 50 μm (right). (E) ddPCR assay of BRAF mutation in primary tumour. BRAFwild, green dots; BRAFV600E, blue dots; no DNA, grey dots. Dots surrounded by red ellipse indicate BRAFV600E.
Table I. Clinical characteristics and BRAF mutations in 9 patients with melanoma
Due to genomic instability, melanoma cells frequently show BRAF mutations and BRAF amplification (5). Because melanoma cells are not necessarily diploid, we cannot simply calculate the proportion of BRAF-mutated cells from the BRAF-mutant AF. At primary tumour sites in case 1, the BRAFV600E AF was low (17.6% and 19.3% at 2 sites), despite the high proportion of melanoma cells (92.1% and 89.2%). Given that melanoma cells were homogeneous, the ratios of BRAFV600E to BRAFwild allele in a tumour cell were calculated to be approximately 1:4.5 and 1:3.9. Furthermore, the BRAFV600E allele was rare (0.02%) in the metastatic lesion. Importantly, it is not uncommon for BRAF AF to differ between the primary and metastatic lesions. Taken together, these results indicate that melanoma cells are more likely to be heterogeneous, rather than homogeneous, in terms of BRAF genotype. The coexistence of BRAF-mutant and BRAF-wild-type tumour cells within the same patient has important implications for clinical decision-making. Based on the tight connection between BRAF genotype and clinical benefits of BRAF inhibitors, it may be necessary to assess BRAF mutations quantitatively. In addition, it is preferable to examine BRAF status in metastatic lesions (if tissue is available) because BRAF status may be different between lesions.
In addition, BRAFV600E can also be detected using a BRAFV600E mutation-specific antibody (6). However, expression of BRAFV600E protein was not detected in a primary lesion of case 1 (data not shown), suggesting that BRAFV600E protein may not be expressed substantially. Further studies are needed to clarify the reason for discordance between BRAF genotype and immunohistochemical analyses.
BRAFV600E is a driver mutation that appears at an early phase of melanoma progression, implying that it is, at least initially, homogeneous (7, 8). By contrast, our results suggest that melanoma is occasionally heterogeneous in the BRAF genotype. Genomic instability produces not only point mutations, but also amplifications and deletions. If melanoma cells harbour multiple aberrant pathways involved in transformation, loss of the BRAFV600E allele may occur without disrupting tumour progression. BRAF heterogeneity was also detected in melanomas using several methods, including single-cell PCR and pyrosequencing (5, 9). Thus, the early appearance of the BRAFV600E allele and the ultimate heterogeneity of the BRAF genotype does not constitute a contradiction.
The BRAFV600E allele was rare in the metastatic lesions of 3 cases we examined (cases 1, 3 and 8), even though the mutant allele was abundant in the corresponding primary tumours. Because metastasis is frequently caused by a tumour cell cluster, rather than a single tumour cell, in peripheral blood (10), the metastatic lesions in these cases may have derived from clusters including BRAFwild melanoma cells along with BRAFV600E cells, and BRAFwild melanoma cells have predominant potential to proliferation and survival. In addition, intra-heterogeneity in the primary lesion should be related to the inter-heterogeneity between the primary and metastatic lesions. However, it is not clear whether a primary lesion with marked intra-heterogeneity results in noticeable discrepancy between lesions.
The authors thank Professor Y. Kawakami of the Keio University School of Medicine, Tokyo, Japan, for providing the Mel-2 melanoma cell line, and Professor T. Nagatani of the Tokyo Medical University, Tokyo, Japan, for providing the A375 melanoma cell line. This study was supported by a Grant-in Aid for Scientific Research No. 15K19685 (to K.S.) from the Japan Society for the Promotion of Science.
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize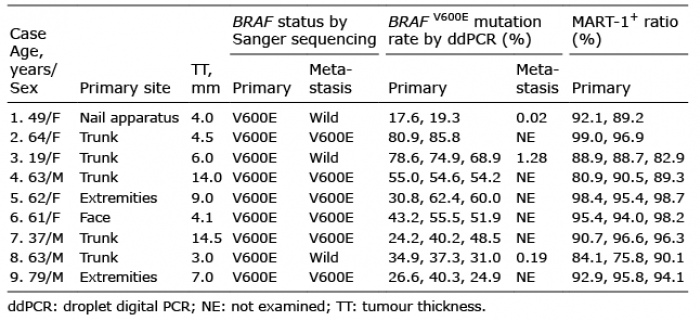 Click to show fullsize
Click to show fullsize