


1Department of Dermatology, University Hospital Muenster, Muenster, Germany, 2Laboratory of Cutaneous Physiopathology, San Gallicano Dermatological Institute, Rome, Italy, 3Biomedical Technology Center of the Medical Faculty, University of Muenster, Germany, and 5Department of Pathology, Fox Chase Cancer Center, Philadelphia, PA, USA
Although recent therapeutic developments raise hope, melanoma remains a devastating disease with a need for new treatment targets. In other tumours prohormone convertases have been shown to be pro-tumourigenic as they are involved in processing preforms of matrix-metalloproteinases, growth factors and adhesion molecules. The aim of this study was to look for new treatment options for melanoma, by investigating the role of the prohormone convertase Paired basic Amino acid-Cleaving Enzyme 4 (PACE4/PCSK6) in melanoma cell lines and human melanoma tissue. PACE4-transfected A375 melanoma cells displayed significantly increased proliferation, MMP-2 production, gelatinase activity and migratory capacity in vitro compared with sham-transfected cells. In vivo, elevated PACE4 expression resulted in significantly increased tumour growth on immunodeficient mice. In the majority of 45 human primary melanomas and melanoma metastases ex vivo PACE4 immunoreactivity was detectable, while it was absent in in situ melanomas. These results indicate PACE4 as a regulator of melanoma cell aggressiveness.
Key words: melanoma; prohormone convertases; carcinogenesis; melanocytes; PACE4; PCSK6.
Accepted May 18, 2020; Epub ahead of print May 25, 2020
Acta Derm Venereol 2020; 100: adv00157.
Corr: Carsten Weishaupt, Department of Dermatology, University Hospital Muenster, Von-Esmarch-Str. 58, DE-48149 Muenster, Germany. E-mail: carsten.weishaupt@ukmuenster.de
Prohormone convertases are enzymes that are involved in regulating the activity of proteins, such as growth factors and adhesion molecules, which are important players in carcinogenesis. Metastatic melanoma is a devastating disease with an urgent need for new treatment options. This study investigated the role of the prohormone convertase PACE4 in melanoma. The results showed that the expression of PACE4 correlates with enhanced proliferation and migratory capacity in vitro and increased tumour growth in vivo. Unlike in situ melanomas it is expressed in advanced human melanomas. Thus, PACE4 is associated with melanoma aggressiveness and may serve as a novel target for melanoma treatment.
Paired basic Amino acid-Cleaving enzyme 4 (PACE4), also designated as proprotein convertase subtilisin/kexin type 6 (PCSK6), is a member of the prohormone convertase (PC) family, which consists of 7 PCs, including furin, PC1/3, PC2, PC4, PC5/6, PC7, subtilisin kexin isoenzyme 1 (SKI-1) and neural apoptosis-regulated convertase-1 (NARC) (1).
As the name implies, PCs are crucially involved in processing of prohormones, such as proopiomelanocortin (POMC) or proinsulin, within neuroendocrine glands. In the skin especially PC1/3 and PC2, the key enzymes processing POMC into adrenocorticotropin, α-melanocyte-stimulating hormone and β-endorphin, have been found to be constitutively expressed in normal human epidermal melanocytes (2), dermal fibroblasts (3) and dermal papilla cells (4). Moreover, PACE4, PC5/6 and PC/8 were found to be expressed by human epidermal keratinocytes in a spatiotemporal manner, indicating a functional role of these enzymes in epidermal proliferation and differentiation (5).
Of note, several lines of evidence indicate that PCs play a role in carcinogenesis (6–8). PCs are involved in processing proteins, such as preforms of matrix-metalloproteinases (MMPs), growth factors, growth factor receptors and adhesion molecules, all of which have an important role in carcinogenesis. Accordingly, increased expression of some PCs, especially furin, PC1, PC2 and PACE4, has been reported in various human cancers and tumour cell lines (9–13). For example, furin expression in tumour cells of head and neck cancer was identified as a marker of aggressiveness (14). Overexpression of furin or PACE4 in distinct tumour cell lines increased their invasiveness in vitro and in vivo, respectively (15–17). Since pharmacological suppression of aberrant PC activity in tumour cells has anti-tumourigenic and anti-metastatic effects, cell-permeable PC inhibitors are currently being discussed for their potential in the treatment of some cancers (18, 19).
However, the role of PCs in human melanoma is still incompletely understood. Thus, this study aimed to investigate the role of PCs in human melanoma. This report focusses on the role of PACE4 in melanoma. The results show that PACE4 can affect biological behaviour of human melanoma cells in vitro and in vivo.
Cells and culture conditions
Melanoma cell lines were cultured in RPMI 1640 supplemented with 10% foetal calf serum (FCS), 1% L-glutamine, 1% penicillin/streptomycin in a humidified atmosphere of 5% CO2 at 37°C. A375 and G361 cells were from ATCC (Rockville, MD, USA) while IPC298, IGR37 and SK-Mel-30 cells were from DSMZ (Braunschweig, Germany). WM35, WM902B and WM9 cells were a gift from M. Herlyn (The Wistar Institute, Philadelphia, PA, USA). WM35, WM902B and IPC298 were established from primary cutaneous melanomas, A375 from a solid melanoma, IGR37, G361, SK-Mel30 and WM9 from melanoma metastases. Normal melanocytes and keratinocytes derived from neonatal foreskin were purchased from Tebu-bio (Portland, USA) and maintained as described previously (20).
Transfection of melanoma cells
A375 melanoma cells were transfected by electroporation with a eukaryotic expression vector containing a 2700-bp wild-type rat PACE4 cDNA fragment (21) or the pCIneo vector only. Two days after transfection, cells were switched to culture medium containing 1 mg/mL G418 (PAA; Cölbe, Germany). G418-resistant colonies were propagated and isolated, followed by confirmation of ectopic rat PACE4 expression using RT-PCR and Western immunoblotting.
Since a human PACE 4 construct was not available and the rat construct had been established through previous work (21) it was decided to use the latter. Comparing human and rat PACE4 nucleotide and protein sequences by using the NCBI Basic Local Alignment Search Tool (22) the human and rat PACE4 transcripts reveal 87% homology and the proteins have 91–97% homology, assuming similar biological activity, which was also proposed by Koide et al. in 2003 (23).
Proliferation assays
Cell proliferation was measured by a commercially available bromodesoxyuridine (BrdU) cell proliferation ELISA (Hölzel Diagnostika, Köln, Germany). A total of 4,000 cells per well were seeded in quintuplicates into 96-well tissue culture plates and incubated with the BrdU labelling solution for 6 h at 37°C, followed by cell lysis, DNA fixation, and incubation with an anti-BrdU peroxidase-coupled antibody. Samples were then incubated in substrate solution, stop solution followed by photometric analysis at 450 nm according to the manufacturer’s protocol.
Determination of melanin content
Measurement of melanin content was performed on 5 × 105 cells seeded in 6-well culture plates. After 24 h cells were detached by trypsinization and centrifuged. Cell pellets were dissolved in 1 M KOH and incubated for 24 h at 37°C in a shaker. Melanin concentration was determined by measuring the optical density of the cell lysates at 405 nm using an enzyme-linked immunoassay (ELISA) plate reader. Melanin content in the samples normalized for protein content was calculated by comparing the sample OD405 with a standard curved generated by synthetic melanin (Sigma, St Luois, MO, USA).
RNA extraction and semi-quantitative RT-PCR
Total RNA was isolated from PACE4 transfectants and controls, normal and transformed melanocytes using a commercial purification kit (Qiagen, Santa Clarita, CA, USA). Primers for detection of rat PACE4 transcripts have been described previously (24). The PCR protocol for rat PACE4 was 1×94°C for 3 min, 30×94°C for 10 s, 60°C for 30 s, and 72°C for 30 s and a final cycle at 72°C for 5 min. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was amplified using the sense primer 5’-TGCACCACCAACTGCTTAGC-3’ and the antisense primer 5’-GGCATGGACTGTGGTC-ATGAG-3’, generating an amplicon of 86 bp by 20×94°C for 10 s, 60°C for 30 s and 72°C for 30 s, followed by a 5-min extension step at 72°C. Amplification products were separated by 1.5% agarose gel electrophoresis and visualized after ethidium bromide staining under ultraviolet C (UVC) light.
Real-time RT-PCR
Quantification of endogenous mRNA levels of PACE4 was determined by real-time fluorescence detection using Absolute SYBR Green ROX Mix (Applied Biosystems, Foster City, CA, USA). For endogenous PACE4 the primer sequences used were sense 5’-TGATTCCTACGCCAGCTACGAC-3’ and anti-sense 5’-TTGTACGCTATGCCCACGATG-3’ generating an amplicon of 153 bp. Primers for 18S rRNA were identical to those published previously (25). Reactions were carried out in an ABI-prism 7300 sequence detector supplied with SDS 2.1 software (Applied Biosystems) using conditions identical to those described previously (20). For comparison of endogenous PACE4 mRNA between melanocytes and melanoma cells the Ct method (ΔCt value) was used.
Western immunoblotting
Western immunoblotting with anti-rat PACE4 (JH1475, 1:1000) directed against the amino acids 570 to 656 of rat PACE4 (26) or anti-human PACE4 (Acris, San Diego, CA, USA, 1:250) directed against the C-terminal region of human PACE4 and anti-MMP-2 (1:2000, Merck, Darmstadt; Germany) was performed according to standard protocols (Appendix S1). The anti-human PACE4 antibody detected an immunoreactive band of ~100 kDa, representing the mature PACE4 (97 kDa) form as well as a weaker band (106 kDa), which resembles the preform of PACE4 (21).
Zymography
Aliquots of conditioned media from PACE4 transfectants and controls were tested for gelatinase activity by separation in a SDS-PAGE 10% gel and staining with Coomassie brilliant blue to visualize bands with gelantinolytic activity (see Appendix S1 for details).
Immunofluorescence staining and immunohistochemistry
Primary cutaneous melanomas and metastases (n = 45) derived from patients undergoing routine surgery were examined. Expression analysis of PCs in melanoma samples was approved by the Local Ethical Committees of the University of Münster (2008-191-f-S). Immunofluorescence and immunohistochemical staining was performed by standard techniques (Appendix S1).
PACE4 gene silencing
A pool of 4 siRNA oligonucleotides (SMARTpool; Thermo Scientific Dharmacon, Bonn, Germany) was used to deplete cells from PACE4. Non-targeting control siRNA was used as negative control. Cells were transfected for 48 h with siRNA at a final concentration of 25 nM following the manufacturer’s instructions.
In vitro invasion assay
A commercially available Biocoat Matrigel invasion assay (BD Pharmingen, San Diego, CA, USA) was used following the manufacturer’s instructions. The tumour number of cells that penetrated through the Matrigel-coated membranes was counted in 3 viewing fields (×100) of 3 independent experiments.
In vitro migration assays using digital holographic microscopy
In order to analyse the migration of the PACE4 transfectants and controls in a 3-dimensional environment a collagen matrix, we performed digital holographic microscopy, as described previously (27–29) (Appendix S1).
In vivo tumorigenesis experiments
SCID Hairless Outbred mice (SHO™) mice were obtained from Charles River, Sulzfeld Germany, bred under specific pathogen-free (SPF) conditions and used at the age of 8–12 weeks. Six mice were injected subcutaneously, either with 1×106 PACE4-transduced A375 cells or with empty vector-transduced melanoma cells. Tumour size was measured 3 times a week with a calliper (Bochem, Weilburg, Germany). Mice were sacrificed when tumour size reached 1 cm3. All experiments were performed according to institutional regulations (8.87-50.10.36.08.286, Office for the Environment, Nature and Municipal Affairs North Rhine-Westfalia, Germany).
Statistical analysis
All experiments were repeated at least 3 times. Descriptive statistical parameters (means ± standard error of the mean (SEM)) were calculated by the SPSS 18.0 software (IBM, Ehring, Germany). Normality of the distribution was checked by the Shapiro–Wilk test, followed by an unpaired double-sided 2-railed Student’s t-test. p-values of < 0.05 were considered significant.
Generation of A375 melanoma cells ectopically expressing rPACE4
In order to investigate whether PACE4 can influence the biological aggressiveness of melanoma cells this study utilized the human melanoma cell line A375, because initial real-time RT-PCR analysis revealed similar relative RNA amounts of endogenous PACE4 compared with normal human melanocytes (NHM) (Fig. 4A). A375 cells were stably transfected with a eukaryotic expression vector encoding rat PACE4 (hereafter called A375.pCI.rPACE4), while control cells were transfected with the vector alone (A375.pCI).
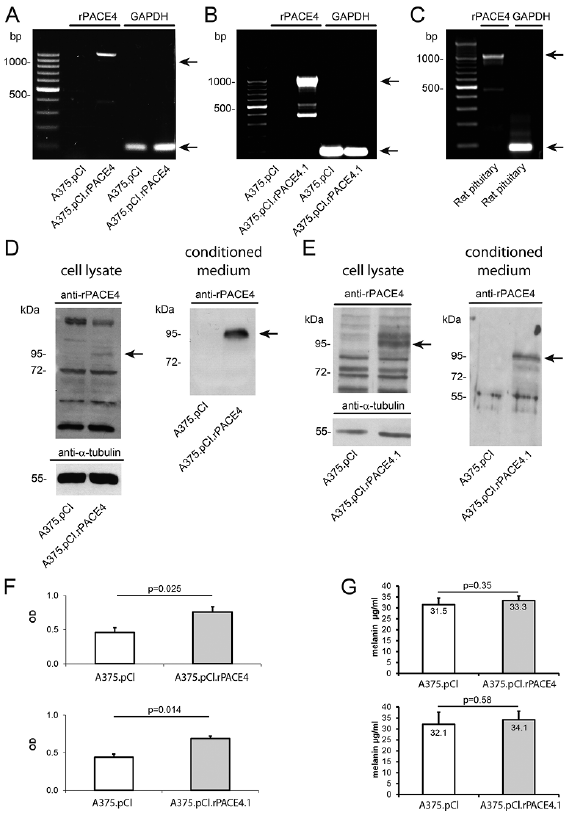
As demonstrated by RT-PCR pooled transfected cells (A375.pCI.rPACE4) and single clone (A375.pCI.rPACE4.1) cells, but not vector-alone-transfected cells, expressed rat PACE4 (rPACE4) (Fig. 1A, B). The PACE4 amplification product of A375.pCI.rPACE4 cells had the same size as the amplicon derived from RT-PCR of rat pituitary gland (1,000 bp) (Fig. 1C). Ectopic expression of rPACE4 in pooled transfected cells (A375.pCI.rPACE4) and single clone (A375.pCI.rPACE4.1) cells was confirmed by western immunoblotting of total cell lysate and conditioned medium. Pooled transfected cells (A375.pCI.rPACE4) and single clones (A375.pCI.rPACE4.1) contained an immunoreactive protein at 97 kDa, which matched the size of mature PACE4 (Fig. 1D, E).
Phase contrast microscopy did not reveal major mor-phological differences between the parental cell line, pooled A375.pCI.rPACE4 transfectants or single-cell clones and A375.pCI cells (data not shown). In vitro proliferation, as measured by bromodeoxyuridine (BrdU) incorporation was significantly increased in both pooled transfected cells (A375.pCI.rPACE4) (p = 0.025) and single clones (A375.pCI.rPACE4.1) (p = 0.014) compared with vector-alone transfected cells (Fig. 1F). This study also assessed the melanin content of the rPACE transfectants and control cells. However, no significant differences could be detected in both pooled transfected cells (A375.pCI.rPACE4) or single clones (A375.pCI.rPACE4.1) compared with vector-alone transfected cells (Fig. 1G).
Fig. 1. Generation and characterization of A375 melanoma cells ectopically expressing rPACE4. (A) Detection of rPACE4 transcripts in pooled clones of A375 transfected with rPACE4 cells (A375.pCI.rPACE4) and (B) the single clone A375.pCI.rPACE4.1, but not in vector-alone transfected cells (A375.pCIneo) as shown by end-point RT-PCR. (C) The rPACE4 amplicon was identical with cDNA obtained from rat pituitary used as positive control. (D, E) Detection of rPACE4 protein in total cell lysates (left-hand panels) and conditioned media (right-hand panels) from pooled clones (A375.pCI.rPACE4) and the single clone A375.pCI.rPACE4.1, but not from A375.pCIneo cells as shown by Western immunoblotting. (F) Increased proliferation of pooled clones of A375 transfected with rPACE4 cells (A375.pCI.rPACE4) and the single clone A375.pCI.rPACE4.1 as measured by BrdU incorporation. (G) Intracellular melanin levels in pooled clones of A375 transfected with rPACE4 cells (A375.pCI.rPACE4) and the single clone A375.pCI.rPACE4.1 as measured by photometric determination. Depicted are representative results from 3 independent experiments (A–F). Data represent means±standard error of the mean (SEM) from 3 independent experiments (F, G), p-value is indicated.
Ectopic expression of PACE4 enhances invasiveness and migration of A375 melanoma in vitro
PCs have been shown to be involved in activation of pro-MMP-2 to its active form by membrane type MMPs (MT1-MMP). Western immunoblotting of conditioned media using an antibody that recognizes activated MMP-2 revealed increased protein levels in A375.pCI.rPACE4 cells compared with vector-alone-transfected cells (Fig. 2A). In addition, gelatine zymography of conditioned media confirmed increased ability of A375.pCI.rPACE4 cells to degrade collagen type IV (Fig. 2B). As these findings may suggest a higher migratory capacity related to PACE4 expression the current study examined the migratory capacity by Matrigel migration assay. Both pooled transfected cells (A375.pCI.rPACE4, p = 0.00000266) and single clones (A375.pCI.rPACE4.1, p = 0.0000287) displayed significantly increased invasiveness, as measured by the Matrigel assay, compared with vector-alone transfected cells (Fig. 2C, D). In addition, digital holographic microscopy (DHM) was employed to track individual melanoma cell movement in a collagen type I gel over time. Here, the single cell clone A375.pCI.rPACE4.1, but not pooled transfectants, displayed increased motility, as measured by DHM over a period of 16 h (Fig. 2E).
Fig. 2. Ectopic expression of rPACE4 in A375 melanoma cells enhances invasiveness and migration in vitro. (A, B) Expression and activity of MMP2 in conditioned media from A375.pCI.rPACE4 and A375.pCIneo cells as measured by Western immunoblot and zymography. (C, D) Pooled clones of A375 transfected with rPACE4 cells (A375.pCI.rPACE4) and the single clone A375.pCI.rPACE4.1 show enhanced penetration through Matrigel-coated membranes; n = 3, **p < 0.001. (E) Analysis of tumour cell migration of the single clone A375.pCI.rPACE4.1 and vector-alone-transfected cells in calf skin type I collagen using time-lapse investigations with DHM. The depicted movement trajectories were obtained from quantitative DHM phase contrast images by tracking the maximum thickness coordinates of selected single cells over an observational time of 16 h. Note, that some, but not all, A375.pCI.rPACE4.1 cells displayed increased migration compared with A375.pCIneo cells. Depicted are representative results from 3 independent experiments.
Ectopic expression of PACE4 in A375 melanoma cells confers increased local tumour growth in immunodeficient mice
As we had observed that ectopic PACE4 expression in A375 melanoma cells was associated with increased proliferation and enhanced proinvasive characteristics in vitro, this study investigated whether this is also relevant for tumour growth in vivo. Analysis of tumour growth of subcutaneously injected cells in immunodeficient SCID hairless outbred (SHO™) mice revealed that A375.pCI.rPACE4.1 cells generated faster growing tumours with enhanced ectopic PACE4 expression compared with empty-vector transfected cells (Fig. 3A, B). The results are depicted as a Kaplan–Meyer survival curve defining a 1-cm3 tumour size, as drop-out survival probability was clearly decreased for PACE4-expressing tumours compared with empty vector expressing tumours (Fig. 3C). Hence, strong expression of PACE4 could also be associated with increased aggressiveness in vivo.
Fig. 3. Ectopic expression of rPACE4 in A375 melanoma cells leads to increased local tumour growth in immunodeficient mice in vivo. 1×106 A375.pCIneo or A375.pCI.rPACE4 cells were injected s. c. into 6 immunodeficient SHO™ mice. Local tumour growth was measured 3 times a week until a maximal tumour size of 1 cm3 was reached. (A) Tumour growth over time as depicted by mean tumour size ± standard error of the mean (SEM); *p < 0.05; n = 3 per treatment group (B) tumour growth of PACE4 and sham-transfected A375 tumours on immuno-deficient SHO™ mice and cytoplasmic detection of PACE protein (green) by immunofluorescence staining in tissue from in vivo grown tumours (magnification 400×). (C) Tumour growth depicted as Kaplan–Meier curve defining a tumour size of 1 cm3 as drop-out.
Endogenous PACE4 is overexpressed in melanoma cells in vitro and ex vivo
Based on the observed growth-promoting effect of ectopic PACE4 expression in A375 melanoma cells in vitro and in vivo this study determined the level of endogenous PACE4 expression in human melanoma cell lines as well as in human melanoma tissue samples. Comparing exponentially growing NHM and 8 melanoma cells lines using real-time RT-PCR analysis, mRNA expression of endogenous PACE4 was significantly increased in 5 out of 8 melanoma cell lines compared with NHM (Fig. 4A). There was no correlation between the endogenous PACE4 mRNA amounts and the pathological tumour thickness of the originating tumour.
At the protein level, endogenous PACE4 expression was assessed in total cell lysates by Western immuno-blotting (Fig. 4B). Interestingly, protein amounts of both PACE4 forms were markedly higher in virtually all tested human melanoma cell lines than in NHM (Fig. 4B). The specificity of the utilized PACE4 antibody was confirmed by gene knock-down using small interference (si)RNA of PACE4. As shown by endpoint RT-PCR analysis transfection of WM902B melanoma cells with PACE4 siRNA did significantly suppress endogenous PACE4 mRNA levels and also reduced immunodetection of PACE4 in total cell lysates using the PACE4 antibody (Fig. 4C, D). In addition, immunofluorescence staining on WM902B cells revealed a clear PACE4 cell membrane staining, as expected, compared with IgG control, although the cytoplasmic background was quite high (Fig. 4E).
Fig. 4. Detection of endogenous PACE4 in human melanoma cells in vitro and in human melanoma tissue samples ex vivo. (A) PACE4 mRNA levels in normal human melanocytes (NHM) and human melanoma cell lines, as shown by real-time RT-PCR. Data represent means ± standard error of the mean (SEM); n = 3; **p <0.01; ***p <0.001. (B) PACE4 protein levels in NHM and human melanoma cell lines, as shown by Western immunoblotting. (C, D) Validation of the specificity of the utilized PACE4 antibody by siRNA. WM902B melanoma cells were treated with PACE4 siRNA or control siRNA vs. the transfection reagent (TR) alone followed by (C) RT-PCR and (D) Western immunoblotting for PACE4 at RNA and protein level. Data are means ± SEM; n = 3; **p <0.01. (E) In vitro immunofluorescence analysis of PACE4 in WM90B melanoma cells. Note: PACE4 immunoreactivity on the cell membrane of WMB90B cells compared with cells incubated with the IgG isotype control. Cytoplasmic staining is unspecific. Space bar: 10 µm.
Finally, PACE4 expression was examined by immunohistochemistry in human melanoma sections derived from 45 patients with melanoma. No PACE4 immunostaining was detectable in normal skin (Fig. 5A,i). Only melanocytes showed low reactivity to anti-PACE4, as it was predicted by low RNA expression of NHM (Fig. 4a). Likewise, all ex vivo in situ melanomas (Clark level I; n = 15) were non-reactive (Fig. 5A,ii). Only a minority (2 out of 11) of early melanomas (Clark levels II (mean tumour thickness 0.35 mm) and III (mean tumour thickness 0.91 mm) displayed weak granular cytoplasmic PACE4 staining within tumour cells.
In contrast, the majority (9 out of 10) of both advanced primary melanomas (Clark levels IV and V (mean tumour thickness 2.59 and 5.83 mm)) and melanoma metastases (8 out of 9) expressed PACE4 immunoreactivity (Fig. 5A,v, vii, ix). PACE4 immunoreactivity in tumour cells of these tissue samples displayed a variable degree of positivity. Incubation of the melanoma sections with the isotype control was consistently negative apart from unspecific staining of melanophages (Fig. 5A, iv, vi, viii, x). While ulceration of the primary tumour did not give a clear correlation to PACE4 expression, a clear positive correlation was noted when PACE4 immunoreactivity was assessed semi-quantitatively dependent on Clark levels/tumour thickness or type of the primary tumour (nodular vs. superficially spreading) (Fig. 5B–D). These findings corroborate the in vitro findings on the impact of PACE4 in melanoma biology which suggested that this enzyme has a growth-promoting and possibly proinvasive effect.
Fig. 5. PACE4 immunoreactivity in melanoma tissue sections. (A) PACE4 immunoreactivity in human melanomas of tissue sections, as shown by immunohistochemistry. (i) normal skin; (ii) in situ melanoma; (iii) Clark level III melanoma with peripheral intraepithelial spreading part (arrows), melanophages (M); (iv) rabbit IgG control of the former, melanophages (M); (v) Clark level IV melanoma; (vi) rabbit IgG control of the former, melanophages (M); (vii) expansion of (v); (viii) rabbit IgG control of the former, melanophages (M); (ix) melanoma metastasis; (x) rabbit IgG control of the former. Scale bar: 100 µm. Degree of PACE4 immunostaining (positivity) of the tumour cells of each tissue section/case was assessed semi-quantitatively by 2 independent investigators using the parameters: absent, weak, strong and very strong. The percentage of tissue sections/cases exhibiting a given staining intensity was then related to (B) Clark level/tumour thickness, (C) type of melanoma (SSM: superficial spreading melanoma; NM; nodular melanoma) or (D) tumour ulceration.
The results of this study show that PACE4 is overexpressed in human primary melanomas and melanoma metastasis. In vitro and in vivo experiments support a tumour-promoting role for PACE4. The molecular mechanism by which ectopic expression of rPACE4 increases cell proliferation of A375 melanoma cells in vitro and enhances tumour growth in vivo remains to be further elucidated. Insulin-like growth factor-1 receptor (IGF-1R) is a crucial PC substrate involved in cell proliferation (30). Blockage of PACE4 activity in murine cutaneous squamous cell carcinoma cell lines with the small-molecule PACE4 inhibitor decanoyl-RVKR-chloromethylketone impaired maturation of IGF-1R, reduced intrinsic tyrosine kinase activity, and decreased cell proliferation in vitro (31). Using a 2-stage skin carcinogenesis model it was further shown that topical administration of decanoyl-RVKR-chloromethylketone reduces tumour incidence, tumour multiplicity and metastasis (31). Mechanistically PACE4 has been reported to exert tumorigenic effects through the mitochondrial pathway and endoplasmic reticulum signalling pathways or an oncogenic alternative splicing switch, resulting in an increase in processing of the pro-growth differentiation factor pro-GDF15 in non-cutaneous tumours (32, 33). Also the cell proliferation- and tumour growth-promoting function of endogenously expressed PACE4 in murine squamous cell carcinoma lines supports our present findings on A375 melanoma cells stably transfected with rPACE4 (17).
Considering the possibility of targeted suppression of PC function in melanoma cells, it was reported that transfection of primary human melanoma M10 cells with the non-selective PC inhibitor α1-antitrypsin (α1-PDX) reduces their in vitro migration and invasion (34). In accordance with the findings from Bassi et al. (31) M10/PDX cells displayed reduced PC activity and were unable to process pro-insulin-like growth factor 1 receptor. Interestingly, α1-PDX was even operative despite altered p53, CDKN2A and N-Ras genes (34). However, as α1-PDX is a non-specific PC inhibitor suppressing the enzyme activities of furin and several other PCs (35), the particular contribution of PACE4 in the upregulation of proinvasive characteristics in M10/PDX cells is unknown. Therefore, in contrast to previous studies, the current study investigated the effects of selective overexpression of rPACE4 in melanoma cells to better delineate the role of PCs in melanoma pathophysiology.
In the context of targeting PCs therapeutically, it should be noted that treatment of human melanoma cells in vitro with the pharmacological subtilisin-kexin-isoenzyme-1 (SKI-1) inhibitor decanoyl-RRLL-chloromethylketone suppressed metabolic activity and induced apoptosis by reduced processing of activating transcription factor 6, a SKI-1 substrate (36). In addition, Levesque et al. had reported on anti-tumour effects of a PACE4 inhibitors in prostate cancer (37). Therefore, it will be of interest to investigate if in the next set of experiments whether suppression of PACE4 by various in vivo approaches also suppresses tumour growth. Such strategies would include PACE4-specific inhibition by small molecules, permanent gene silencing, e.g. by the Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR/Cas) methodology. Our immunohistochemical finding that PACE4 immunoreactivity is undetectable in melanoma in situ, but present in the majority of advanced primary melanomas (Clark levels IV and V, mean tumour thickness 2.59 mm/5.83 mm) and metastases supports the concept of an upregulatory role of endogenous PACE4 on cell proliferation and tumour growth. Further studies on a larger cohort of melanoma tissue sections are needed to clarify whether PACE4 can be used as a prognostic marker for melanoma independent of tumour depth.
In summary, this study shows that PACE4 is a novel player in the pathobiology of melanoma. Targeting PACE4 may be an interesting novel strategy in the future therapy of melanoma.
The authors thank Gert von Bally for scientific support, Ilka Wolff, Britta Ringelkamp, Andrea Wissel, Angelika Vollmer and Steffi Ketelhut for expert technical assistance. The work was supported by the Innovative Medizinische Forschung of the University of Münster (grant number: WE110817) to CW and MB and by the Germany Federal Ministry for Education and Research to BK and GvB (grant number: FKZ 13N10937).
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize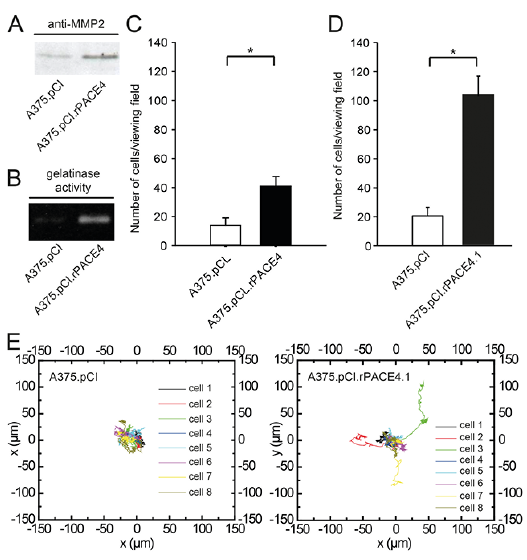 Click to show fullsize
Click to show fullsize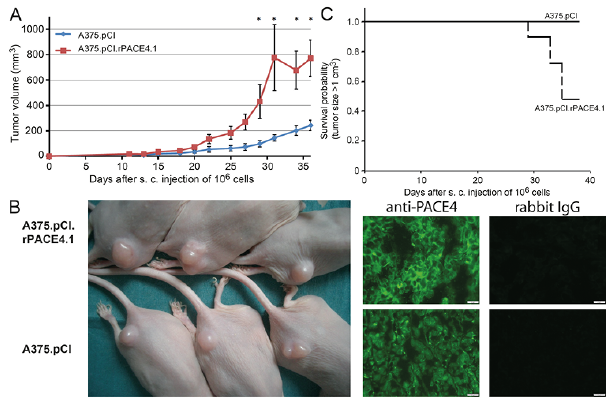 Click to show fullsize
Click to show fullsize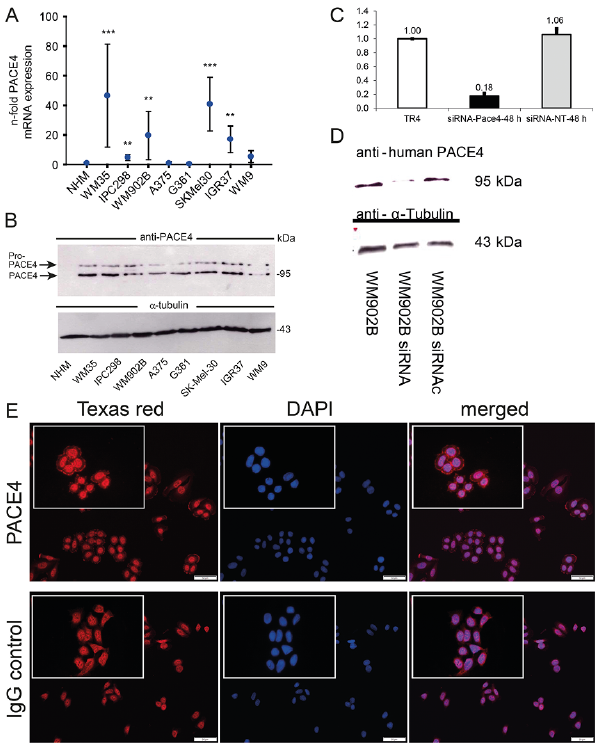 Click to show fullsize
Click to show fullsize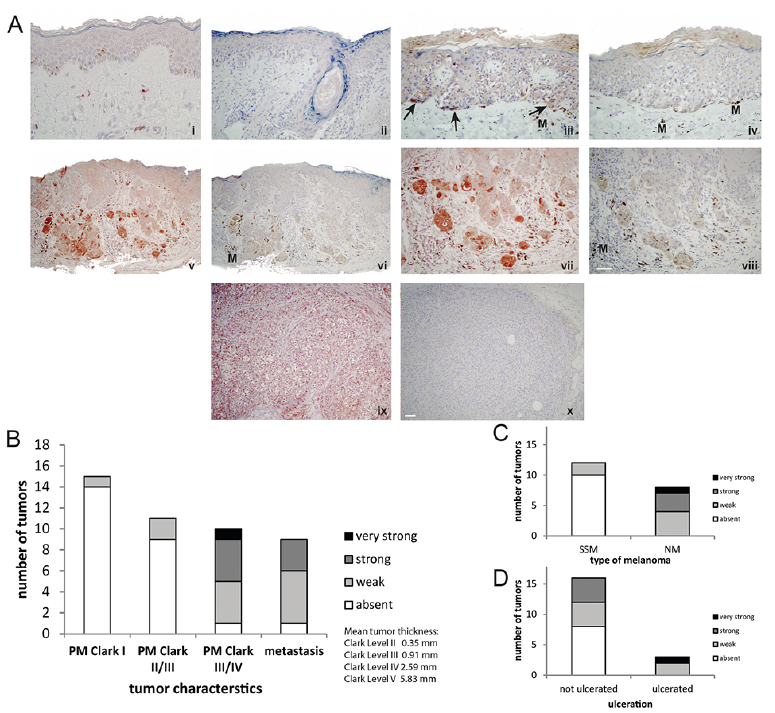 Click to show fullsize
Click to show fullsize