


Darier disease is a severe, rare autosomal dominant inherited skin condition caused by mutations in the ATP2A2 gene encoding sarcoendoplasmic reticulum Ca2+-ATPase isoform 2 in the endoplasmic reticulum. Since sarcoendoplasmic reticulum Ca2+-ATPase isoform 2 is expressed in most tissues, and intracellular calcium homeostasis is of fundamental importance, it is conceivable that other organs besides the skin may be involved in Darier disease. This review focusses on the association of Darier disease with other organ dysfunctions and diseases, emphasizing their common molecular pathology. In conclusion, Darier disease should be considered a systemic condition that requires systemic and disease mechanism targeted treatments.
Key words: Darier disease; genodermatosis; rare disease; endoplasmic reticulum; calcium; SERCA2.
Accepted Feb 16, 2021; Epub ahead of print Feb 19, 2021
Acta Derm Venereol 2021; 101: adv00430.
doi: 10.2340/00015555-3770
Corr: Etty Bachar-Wikström and Jakob D. Wikström, Dermato-Venereology Clinic, Karolinska University Hospital, SE-171 64 Stockholm, Sweden. E-mails: ester.bachar-wikstrom@ki.se; jakob.wikstrom@ki.se
While research on the systemic aspects of Darier disease is still in its infancy, it is becoming clear that Darier disease is a multi-organ systemic condition. This not surprising, considering the overwhelming experimental evidence of the importance of sarcoendoplasmic reticulum Ca2+-ATPase isoform 2 in physiology and pathophysiology. While more data is needed, treating physicians should be aware of the risk of extracutaneous manifestations of Darier disease. Future research would benefit from more registry studies, as well as from systemic large-scale Darier disease cohort studies, in order to examine associations as well as to find direct experimental evidence of other organ dysfunctions and diseases.
Darier (Darier-White) disease (DD), also known as keratosis follicularis, is an autosomal dominant inherited skin disease, which is thought to be caused by mutations in the ATP2A2 gene encoding sarcoendoplasmic reticulum Ca2+-ATPase isoform 2 (SERCA2) (1). The disease was described independently by Darier and White in 1889, and the estimated prevalence ranges from 1 in 30,000 people in Scotland and 1 in 36,000 people in northern England, to 1 in 100,000 people in Scandinavia (2–4).
Since cellular calcium homeostasis is of fundamental importance, and the causative mutations are endogenously expressed, it is not surprising that a growing body of evidence indicates that DD is associated with non-dermatological conditions, such as epilepsy, neuropsychiatric disorders including mild intellectual disability, psychiatric disorders such as bipolar disorder and schizophrenia, as well as diabetes (5) and heart failure (6). This paper reviews DD and its association with the skin and other organs, emphasizing the connection between the molecular pathology that underlies these associated common conditions and DD.
Mutations in the ATP2A2 gene were shown to be associated with DD in a landmark study in 1999 by Hovnanian et al. (1), which examined 13 families and sporadic cases. In an earlier publication, Parfitt et al. (7) mapped the disease genes to chromosome 12q23-q24.1, and the specific loci, which are between D12S78 and D12S79. The inheritance of one faulty copy of the gene causes a decrease in transcript production, which causes disease (haploinsufficiency) (8). Subsequently, more than 270 unique ATP2A2 mutations that disrupt critical functional domains of SERCA2 have been associated with DD (8, 9). These mutations vary between families and do not show gene clustering meaning (10). Most mutations described are missense mutations, in-frame deletions or insertions (63%) (http://www.hgmd.org). Other mutations will cause premature termination codons (PTC) or aberrant splicing (37%). Several studies aimed at identifying mutations leading to SERCA2 dysfunction or reduced expression, used a PCR amplification of exons and adjacent intronic splice sites of ATP2A2 (1, 11, 12). Thus, intronic mutations could have been missed. In an approach with similar coverage, Leong et al. used whole exome sequencing (WES) to screen the ATP2A2 gene in patients with DD, and identified 15 novel variants that were not previously reported in DD; however, when genotype-phenotype association was examined for all variants in relation to the patient’s disease severity score, no correlation could be established (13, 14). On the other hand, missense mutations were reported to be associated with a more severe disease phenotype elsewhere (1). Of note, out of 28 patients examined by Leong et al. (13), 4 patients did not have any variants in the ATP2A2 gene despite being clinically diagnosed with DD, and 2 patients had benign ATP2A2 variants. Although examined DD cohorts are small, the percentage of detected mutations is similar; Ringpfeil et al. (15) (58%), Onozuka et al. (16) (70%), and Leong et al. (13) (57%) . Thus, while the majority of DD cases appear to associate with ATP2A2, it cannot be excluded that undetected intronic mutations or deletions in this or even other genes could play a role in a subset of patients. Whole genome sequencing of DD cohorts would be needed to study this further.
Skin symptoms and clinical features
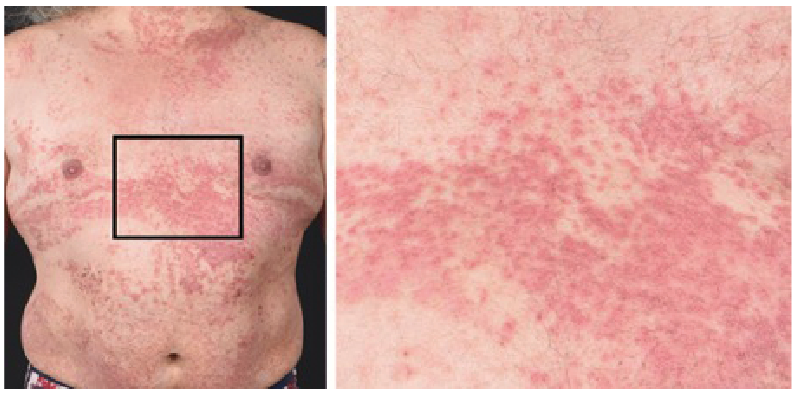
DD is characterized primarily by malodorous warty, greasy, yellow to brown, hyperkeratotic papules, on the seborrheic areas of the chest, upper back, forehead, scalp, nasolabial folds, and ears (Fig. 1) (2, 17). These lesions can lead to large crusted plaques with a high risk of acquiring secondary infections, particularly in folded areas (18). Typical nail abnormalities are characterized by longitudinal white or red lines with ridges and distal v-shaped notches on the nail surface (19, 20). In addition, papules may appear on mucosal membranes, mainly oral, pharynx, vulva and rectum (18). The whitish oral mucosal lesions mostly affect the hard palate and resemble nicotinic stomatitis (21). The gingivae is also a very commonly affected intraoral site, and when the disease affects the tongue it gives rise to a coarse, nodular appearance to the dorsum (22). In the majority of cases, the disease develops in adolescents or young adults and continues throughout life. Ultraviolet B (UVB) irradiation, heat, friction, and infections of affected areas are clinically known to exacerbate symptoms. The impact of DD goes beyond physical symptoms as shown by a severe reduction in health-related quality of life, which correlates with various disease characteristics, such as the skin area involved (23). The treatment of DD can be challenging and is often unsatisfactory (19). Currently, the most effective treatment is systemic retinoids, which may reduce hyperkeratosis, although the primary disease mechanism is not targeted.
Fig. 1. Patient with typical Darier disease, with cutaneous red papules that coalesce into plaques. Black square: inset magnified area. Patient consent for publication was given.
Histopathological features
Microscopic examination of DD lesions reveals hyperkeratosis and hypergranulosis along with central keratin plugs. The majority of DD biopsies exhibit suprabasal clefts with acantholysis that extends through all levels of the epidermis with papillated epidermal hyperplasia (24–26). Examination of the epithelium reveals varying numbers of dyskeratotic cells in the stratum spinosum and stratum granulosum, described as “corps ronds” (round bodies) and parakeratotic cells in the stratum corneum which resemble “corps grains”. The underlying dermal connective tissue is fibrocellular and with mild chronic inflammation (25, 27), although in a case report neither neutrophilic nor eosinophilic infiltrates were observed (24). DD diagnosis is still primarily made with histopathology; however, it is expected that genetic tests will complement or replace this.
Keratinocyte adhesion
The epidermis of the skin is a thin, yet strong, protective layer. One of the classical histopathological signs of DD in skin biopsies is reduction in epidermal cell adhesion, acantholysis, mediated by a rupture of the desmosome-keratin complex (28). Desmosomes as well as adherens junctions are 2 types of cell-cell junctions with specific anchorage to keratinocyte intracellular intermediate or actin filaments that allow strong adhesion between cells and provide the epidermis with strength and resistance to mechanical stress (29, 30). The importance of Ca2+ homeostasis for the integrity and function of the epidermis has been studied extensively. For example, Duden & Franke (31) showed that keratinocytes cultured in media with low concentrations of Ca2+ show desmosomes disassembly, while physiological Ca2+ concentrations in the media direct desmosomal proteins to the plasma membrane where they form mature desmosomes (32). In DD patient-derived keratinocytes, as well as in keratinocytes treated with pharmacological inhibitors of the SERCA pumps, impairment of trafficking and distribution of desmosome components including desmoplakins, desmoglein, and desmocollin was found (33, 34). This may be due to extracellular increase in Ca2+, as this was shown to trigger transient desmosomal remodelling (35), or ER stress, as the molecular chaperone miglustat rescued ER stress impairment of cell-cell adhesion (36), although these are probably connected as ER stress is often associated with altered Ca2+ homeostasis. Li et al. (37) showed that, in human keratinocytes, pharmacological inhibition of SERCA impairs post ER maturation of desmosomal cadherins (DC), which, in patients with DD are mislocalized, further demonstrating that ER Ca2+ homeostasis is crucial to the ER to Golgi transport of nascent DC and that its impairment compromises keratinocyte adhesion. Similar results were shown in a canine kidney cell model for intercellular junction assembly, where inhibition of SERCA led to attenuating the formation of tight junctions and desmosome connections (38). It would be interesting to examine whether dysfunction in other ER Ca2+ regulators besides SERCA2 could create DD-like phenotypes.
Dyskeratinization and calcium dyshomeostasis
The epidermis continuously self-renews as keratinocytes grow outwards and differentiate. The gradient of calcium across the epidermis plays a crucial role in the keratinocyte differentiation, which is evident by simple cell culture experiments in which keratinocytes switch from proliferation to differentiation when cultured in high [Ca2+] (39). Recent studies implicate the ER as the major Ca2+ storage compartment, which forms the epidermal Ca2+ gradient (40–43). Indeed, histopathological examination of biopsies from patients with DD show focal dyskeratosis (premature differentiation of single keratinocytes) and hyperkeratosis (28). The round dyskeratotic cells termed “corps ronds” in the stratum spinosum and “grains” in the upper layers of the epidermis are probably apoptotic keratinocytes (28, 44). Leinonen et al. found lower epidermal Ca2+ in DD lesional compared with non-lesional skin (45); probably, however, not shown due to less SERCA2 expression and perhaps also due to skin barrier disruption, as this leads to ER Ca2+ release to the extracellular space (35).
Importantly, the majority of ATP2A2 mutations associated with DD exhibit a reduced expression and activity of the SERCA2 Ca2+ pump (46–50), therefore, an imbalance in cellular Ca2+ signalling is a hallmark in DD. SERCA2 is a 110-kDa transmembrane protein located in the ER, which is responsible for transporting Ca2+ back into the lumen of the ER from the cytosol (51) (Fig. 2). The overall structure is comprised of 3 cytoplasmic domains: an actuator, a nucleotide-binding, and a phosphorylation domain, as well as 10 transmembrane helices (52). Several studies show that SERCA is activated after binding of 2 Ca2+ ions, a process, which is ATP dependent and forms a phosphoenzyme, followed by release of Ca2+ ions into the ER/SR (53). Over the years, several studies, both in humans and in a murine model of the disease (heterozygous mice), suggested possible mechanisms leading to SERCA2 dysfunction. Ahn et al. (49) showed that most mutations in SERCA2b (out of 12 tested in this study) markedly affected protein expression; partially because of enhanced proteasomal degradation of mutant protein. Moreover, there was also a dominant negative effect of mutants over wild type (WT) pumps (49). In a murine DD model, Zhao et al. (54) showed that amylase exocytosis was ≈10-fold more sensitive to Ca2+ in cells from SERCA2+/– mice compared with WT-derived cells, which suggests a plasticity and adaptability of Ca2+ signalling and Ca2+-dependent cellular functions in vivo, which can explain the relatively normal function of most cells in patients with DD and that heterozygosity is compatible with life. Important to note is that SERCA2, specifically the SERCA2b isoform, is the SERCA type with the highest Km (55) and thus the highest Ca2+ pumping capacity, which may explain the vast impact of its mutations and why mutations in SERCA1 or SERCA3 have not been implicated in DD. SERCA pumps are encoded by 3 genes in humans (ATP2A1-3) that generate multiple isoforms (SERCAla,b, SERCA2a-c, SECA3a-f) by developmental or tissue-specific alternative splicing (56). These pumps differ by their regulatory and kinetic properties, allowing for optimized function in the tissue where they are expressed.
Fig. 2. (A) Endoplasmic reticulum in a human keratinocyte as seen by transmission electron microscopy. ER: endoplasmic reticulum; M: mitochondrion; K: keratin; N: nucleus. (B) Principles of calcium homeostasis in the endoplasmic reticulum. SERCA1-3 pumps Ca2+ into the ER, while the channels ryanodine receptor (RyR) and inositol trisphosphate receptor (IP3R) releases Ca2+ from the ER. The Ca2+ concentration is substantially higher in the ER than in the cytosol. Adapted from (182).
Endoplasmic stress
ER stress is defined as an imbalance between the protein load and the folding capacity of the ER, which results in accumulation of misfolded proteins, further activating an adaptive response termed the unfolded protein response (UPR). In mammals, The UPR response is carried out by 3 ER stress transducers, namely PKR-like endoplasmic reticulum kinase (PERK), inositol-requiring 1α (IRE1α), and activating transcription factor 6 (ATF6). The homeostatic impact of the UPR is played out at several levels (57). First, cells focus on translation attenuation to reduce the ER protein load. Secondly, the UPR upregulates the folding machinery by inducing ER chaperone genes. Thirdly, the ER compartment expands to accommodate the high protein load, and then activates ER-associated degradation (ERAD) of unfolded or misfolded proteins. However, if the UPR fails to resolve prolonged ER stress, apoptotic signals appear and cell death occurs. Thus, there exists a therapeutic window for balancing ER stress before tissue damage occurs.
ER stress is usually associated with ER calcium dyshomeostasis and the SERCA inhibitor thapsigargin is widely used to elicit ER stress experimentally. An immunohistochemical analysis of several ER stress markers in skin lesions derived from a patient with DD suggested a role of ER stress in the pathogenesis of DD (16). A vast genetic analysis of diverse mutants identical to those found in patients with DD demonstrated that SERCA2 mutated protein itself can initiate ER stress (51). These SERCA2 mutated proteins were found to be less soluble, to aggregate, and to be more polyubiquitinylated. After transduction into primary human keratinocytes, mutated SERCA2 aggregates elicited ER stress, led to increased numbers of cells to round up and detach from the culture plate, and induced apoptosis (51). Also, keratinocytes isolated from patients with DD exhibit prolonged ER stress, specifically, increased phosphorylation of eIF2α and IRE1α, which further contributes to aberrant cell-cell adhesion and keratinization and stimulates apoptosis (33, 58), while treatment with the drug miglustat restores adherens junctions and desmosomes in DD keratinocytes (33). One suggested explanation to the miglustat rescue effect is its action as a chaperone that allows the adhesion molecules to evade the UPR, reach the plasma membrane and form adhesion junctions and desmosomes. Another explanation offered by the authors is that miglustat, which is used clinically for type I Gaucher disease, also inhibits glucosylceramide synthase (59) and thus may act through its modulation of the ceramide/sphingolipid pathway (60).
Although speculative, it may be that ER stress induces preterm keratinocyte apoptosis that is identified histopathologically as corps rounds.
Diabetes types
Diabetes refers to a group of metabolic diseases characterized by hyperglycaemia resulting from defects of insulin secretion and/ or increased resistance to insulin. Type 1 diabetes is developed following the destruction of β-cells in the pancreas, which leads to absolute insulin deficiency and full dependency on exogenous insulin for treatment. There are 2 main forms of type 1 diabetes; one is an immune-mediated disease with autoimmune markers, while the second form of type 1 diabetes named idiopathic diabetes, has no known cause and only a minority of patients fall into this group. Type 2 diabetes is by far the most common form of diabetes. In type 2 diabetes, the response to insulin is diminished, defined as insulin resistance (61). The disease is most commonly seen in people older than 45 years, but is increasingly seen in children, adolescents, and younger adults due to rising levels of obesity, physical inactivity, and high-calorie diets (62, 63).
Insulin secretion
In the pancreatic β-cell, glucose is acting as the major stimulator of insulin release. When the concentration of glucose increases, β-cell metabolism accelerates, leading to an increase in ATP/ADP ratio, which induces closure of KATP channels (64, 65). The resulting decrease in K+ efflux causes plasma membrane depolarization, followed by opening of Ca2+ channels and Ca2+ influx into the cell. The subsequent increase in cytosolic Ca2+ concentrations then promotes exocytosis of insulin granules (66, 67). The triggering Ca2+ signal is crucial. Experimental conditions that hamper the increase in cytosolic Ca2+ concentrations impair glucose-induced insulin secretion, while physiological or pharmacological means that increased β-cell cytosolic Ca2+ concentrations induce insulin secretion (68, 69).
In β-cells, as well as other cell types, there are 2 main Ca2+ release channels in the ER; the ryanodine receptor (RyR) and the inositol trisphosphate receptor (IP3R), which controls Ca2+ efflux (70, 71). SERCA2b and SERCA3a are the main SERCA isoforms found in the pancreatic β-cells (72) and control ER Ca2+ influx. These pumps ensure glucose-stimulated insulin release from the β-cell by maintaining high Ca2+ levels in the ER via pumping Ca2+ from the cytoplasm into ER. The involvement of SERCA2b/3a, as well as reduced RyR expression or function, in the pathogenesis of type 2 diabetes was reported (31, 73, 74). Furthermore, the role of ER stress in diabetes development has been massively investigated due to the fact that pancreatic β-cells are exposed to marked changes of ER protein load in response to daily physiological changes in insulin demand. The pancreatic β-cells are therefore, highly susceptible to ER stress under conditions of increased insulin production, as occurs during prolonged hyperglycaemia (75, 76). Chronic ER stress associated with diabetes can impair protein folding in the ER, reduce insulin secretion, induce oxidative stress, and lead to β-cell death (77). In this context, ER stress might occur due to pathological, environmental, and genetic factors including glucolipotoxicity, inflammatory responses, amyloid accumulation, and expression of mutant proinsulin (78–81). The indication that ER stress may play a role in the pathogenesis of autoimmune diabetes came from recent reports that demonstrate the presence of some of the ER stress markers in inflamed islets of both diabetes-prone non-obese diabetic (NOD) mice (82) and patients with autoimmune diabetes (83). Engin et al. (84) showed diminished expression of the UPR markers spliced XBP-1 and ATF6 in β-cells of NOD mice and human patients during the disease progression. This study emphasizes the delicate balance of the unfolded protein response to ER stress, as was demonstrated previously (85). A continuous diabetic environment might lead to prolonged ER stress overcoming the cell capability to reduce ER stress by failing to activate UPR branches. In this case, a failed adaptive UPR response can correlate with β-cell death and insulin deficiency in both experimental models and human T1D (84).
Due to the importance of ER calcium homeostasis for β-cell physiology and pathophysiology, Cederlöf et al. (86) examined the potential association of DD with diabetes at the population level. Individuals with DD had a 74% increased risk of being diagnosed with type 1 diabetes; however, no increased risk of type 2 diabetes was found. This study was performed in a Swedish population, and more data acquired from other populations will be needed to verify DD as a risk factor for type 1 diabetes. Several monogenic forms of diabetes provide strong evidence for the crucial role of single molecules in causing diabetes in humans. For example, INS1 mutation or the Akita insulin mutation (the later initially described in mice (87)), will lead to neonatal diabetes (88), maturity onset diabetes of the young (MODY) or antibody negative T1D. In addition, mutations in EIF2AK3 (part of the PERK signalling arm of the UPR) cause Wollcot-Rallison syndrome (89), and recessive mutations in WFS1, encoding the ER resident protein wolframin 1 that regulates calcium and ER protein folding, leads to Wolfram syndrome 1. Of note, some dominant mutations in WFS1 can also cause Wolfram syndrome-like disease, which can appear with or without diabetes (90–92). Both of these autosomal recessive disorders show ER stress and cause young-onset diabetes (93). In accordance with this, several studies show the importance of proper functioning SERCA2 in preventing β-cell death and diabetes in response to ER stress (35, 94, 95). A recent study supports an important role of SERCA2 in β-cells in humans, as patients with DD have an altered β-cell phenotype (5). Specifically, evidence of basal hyper-insulin secretion was found by the Homeostasis Model Assessment HOMA2-%, a marker for basal insulin secretion (5). It is notable that many more genes, mainly Human Leukocyte Antigen (HLA)-related, are linked to different forms of diabetes, as described in detail by Yang et al. (96).
What are the possible molecular causes behind type 1 diabetes risk in DD? Several of the autoantigens in autoimmune diseases, such as coeliac disease (97), collagen-induced arthritis (98), multiple sclerosis/experimental autoimmune encephalomyelitis (99), rheumatoid arthritis (100), and systemic lupus erythematosus (101) undergo post-translational modifications in the ER, which is important for the proteins’ normal function, as well as antigen properties. ER stress per se can also activate enzymes that alter post-translational modifications of proteins and thus create neoautoantigens, which have a pathological role in type 1 diabetes (102). Likewise, some of the environmental triggers for type 1 diabetes, such as Coxsackie viral infection, have been shown to increase β-cell ER stress, which may increase production of autoantigens or create neo-autoantigens (103). Autoantigens are sensed by dendritic cells engulfing apoptotic β-cells and stimulate the maturation of β-cell reactive T-cells. Hence, abnormal ER stress may stimulate immuno-destruction of β-cells. DD and type 1 diabetes typically both debut in adolescence, which suggests a possible common pathophysiology. While speculative, it may be that ER stress in β-cells in patients with DD interferes with normal protein folding and thus creates neoautoantigens in a similar manner, which induce regular autoimmune β-cell destruction.
Cardiomyocyte calcium homeostasis
The 2 major SERCA2 protein isoforms are the housekeeping SERCA2b, which is expressed in all tissues at low levels (104), and the more specialized SERCA2a isoform, predominantly expressed in cardiac and slow-twitch skeletal muscle (105). In the heart, a raised intracellular Ca2+ concentration is the trigger that activates cardiomyocyte contraction. Specifically, Ca2+ enters through the L-type channels, located primarily at sarcoplasmic reticulum (SR) junctions. The influx of Ca2+ triggers the release of further Ca2+ from the SR via ryanodine receptor 2 (RyR2). Elevated free intracellular Ca2+ will result in interaction between actin and myosin, shortening of sarcomeres and contraction. Diastolic relaxation is an active (ATP-dependent) process, in which Ca2+ transport out of the cytosol occurs via the SR Ca2+ ATPase SERCA2a, leading to a decrease in intracellular Ca2+ concentrations required for both muscle relaxation (106) and for replenishing Ca2+ stores needed for the next contraction (107).
It is widely described that deranged cardiomyocyte Ca2+ kinetics can cause heart failure (108). Heart failure (HF) is a major public health problem, associated with significant mortality, morbidity, and healthcare expenses, particularly among those above the age of 65 years (109, 110). SERCA2a expression is downregulated in heart failure (111–114). Consequently, in vivo gene transfer of SERCA2a in pigs preserved systolic function and improved ventricular remodelling (115), whereas restoring SERCA2a expression by gene transfer in heart-failure patients showed some contradicting results (116, 117). Thus, more research is needed into the significance of SERCA2 in human heart failure.
An important common pathological event between HF and DD is ER stress. As mentioned earlier, depletion of ER luminal Ca2+ may induce the UPR. UPR-induced apoptosis has been implicated in the pathophysiology of heart failure, as well as in other cardiovascular diseases (118–123). Specifically, patients with HF display structural and architecture alteration of the ER, as well as maladaptation of the ER proteins involved in the UPR (124). Several ER stress markers, such as spliced X-box binding protein 1 (XBP1s), glucose-regulated protein 78 (GRP78), activating transcription factor 4 (ATF4), and C/EBP-homologous protein (CHOP) were all shown to be induced in HF in humans (118, 124–127). In conjunction with the human data, animal studies, mainly in mice, showed a protective role of ER stress in HF; hearts of PERK knockout mice showed a significant reduction in Serca2α expression, an increase in apoptosis and UPR genes expression (GRP78, GRP94, CHOP) in response to induction of heart failure (128), and thrombospondin (Thbs) knockout mice showed reduced activation of Atf6α with injury and Thbs4-mediated protection was lost upon Atf6α deletion (129). Overall, the involvement of ER stress in HF is well established, however, one should bear in mind the differences between the models studied and the implications for humans.
Since DD and HF display a common pathological mechanism due to perturbation in SERCA2, the susceptibility of patients with DD to HF was further examined. Interestingly, SERCA2 haplosufficient mice were reported to develop HF when crossed with a transgenic model of increased myofibrillar Ca2+-sensitivity, suggesting that patients with DD might be more susceptible to HF (8). Indeed, in a recent publication, it was shown that patients with DD exhibit a disease-specific increased risk of HF at an earlier age compared with healthy controls (6). Interestingly, female patients with DD showed a higher risk for heart disease in general. This observation contributes to the evidence of an important role of SERCA2 in human HF pathophysiology and suggests that perhaps HF in patients with DD is a specific subtype that requires tailored treatment. More studies are needed to corroborate DD as a risk factor for HF.
The idea that patients with skin conditions (e.g. psoriasis, eczema, and skin cancer) frequently face psychological challenges, such as depression, anxiety and suicidality and overall poor quality of life (130–132) is well known. This is attributed to the stress related to the avoidance-coping mechanism, as well as social and activity prevention. However, in the case of patients with DD, the link between the severe skin condition and psychological problems is likely to be of a genotype-phenotype correlation in many cases and, in fact, DD-causing mutations in ATP2A2 increase the susceptibility to neuropsychiatric dysfunction, in particular severe psychiatric illness (133). The mechanism underlying this correlation probably lies in the important role of Ca2+ in brain function. Indeed, disruption of normal Ca2+ flux in the brain has been associated with psychiatric conditions, including bipolar disorder, schizophrenia, autism spectrum disorders, and intellectual disabilities (134, 135), as well as neurodegenerative disorders, such as Alzheimer’s disease (136, 137) and Parkinson’s disease (138). As in the case of heart and skin, in the neuronal system, Ca2+ released into the cytoplasm is pumped back into the ER by SERCA2 (139). SERCA2a expression has been identified in the brain at low levels, and is found in the granular cells of cerebellar Purkinje neurones, as well as in the giant cells of the brainstem reticular formation (140, 141). SERCA2b is expressed at higher levels in neuronal microsomes, synaptic plasma membrane vesicles, and synaptosomes (142). An early association between SERCA2 and neuropsychiatric pathophysiology was made in patients with DD by Jacobsen et al. (135) and by Craddock et al. (143), and additional work has shown that patients with DD have an increased prevalence of several neuropsychiatric disorders, including depression (30%), bipolar disorder (4%), epilepsy (3%), schizophrenia (1%) and cognitive disabilities (4%) (31, 144–148). One possible explanation for common effects of mutations in the SERCA2 gene on skin and brain is that these tissues share a common ectodermal origin and thus may share sensitivity towards SERCA2 impairment (149). Interestingly, the drug lithium, which is used to treat bipolar disease and acts to alter intraneuronal Ca2+ concentrations, distribution and signalling (150, 151), was reported to exacerbate or even cause DD (152, 153) and, indeed, lithium was shown to decrease SERCA2 expression in rats (154). It would certainly be interesting to examine whether specific ER Ca2+ targeting treatments would impact the different neuropsychatric conditions associated with DD. While the neuropsychiatric association of DD is well explored, further research is needed to examine whether DD is associated with neurodegenerative conditions.
It is not known whether DD is associated with cancer; however, several in vitro and animal studies imply that this is worth examining. An ageing study in heterozygous Atp2a2–/+ mice revealed a higher incidence of squamous cell carcinoma of the oral mucosa, i.e. of keratinocytes, which are also affected in human DD (155). Human-derived primary oral tumours showed that the ATP2A2 gene is downregulated, which may be partly regulated by an epigenetic mechanism (156). Moreover, downregulation of both SERCA2 and SERCA3 expression was reported in thyroid and colon cancer cell lines (157, 158). Korosec et al. (159) showed that germline alterations of ATP2A2 in humans may predispose to lung and colon cancer, and that an impaired ATP2A2 gene might be involved as an early event in carcinogenesis. Mechanistically, studies in mice showed that Atp2a2 haploinsufficiency dramatically affects keratinocytes overall gene expression and differentiation, leading to alteration of the tissue environment, which may be permissive for tumour development (160, 161). In addition, biopsies from patients with colorectal cancer showed high expression of SERCA2 (162) and overexpression of SERCA2b in human colon adenocarcinoma (162), and liposarcoma (163) promoted cell proliferation and migration. Thus, SERCA2 mutations, such as in DD, may affect oncogenesis at several levels. Although speculative, it may be that DD is associated with increased risk for some malignancies, and reduced risk for others.
As for many other rare diseases, a long list of suggested treatments usually indicates that no one treatment works well. Treatments for mild symptoms of DD include moisturizers, sun protection, and careful selection of clothing to avoid heat and sweating (20). Localized DD lesions can be treated with topical glucocorticosteroids or vitamin D3 ointment (164), mainly aiming to reduce inflammation without altering the disease course (165). Several topical retinoids, such as isotretinoin (166), tazarotene (167), and adapalene (168), were also reported as DD treatments. Topical retinoids and retinoid analogues help normalize cell turnover and cell cohesion (169), with isotretinoin also exhibiting a strong inhibition of cell proliferation (170, 171). Ultimately these actions facilitate the levelling of papules and reduce hyperkeratosis. It is not known if retinoids have any effect on keratinocyte calcium homeostasis, thus targeting the specific DD pathology, although in ovarian follicle granulosa cells retinoic acid was shown to increase ER stores of Ca2+ (172), and in myelogenous and promyelocytic bone marrow cells retinoic receptor activation was associated with reduced ER stores of Ca2+ (173). Moreover, in vitro studies have demonstrated an anti-inflammatory effect of retinoids, which might be of additional benefit in DD, since the skin lesions can become inflamed (174, 175). For effective removal of DD refractory-proliferative lesions surgical approaches, such as dermabrasion, electrodessication, and various ablative laser modalities, have sometimes been used (176). In clinical routine, severe symptoms of DD are currently mainly treated systemically with the oral retinoids acitretin (177) or isotretinoin (178). Although showing efficacy on skin lesions (170), recurrence always occurs after ceasing treatment and adverse effects are common (170). These include dry lips, cheilitis, scaling, skin atrophy and fragility, as well as hepatotoxicity, ophthalmological complications, pancreatitis, and skeletal alterations (179). Consequently, patients might undergo intermittent treatment or even discontinuation of treatment (4). Of note, in rare cases DD has also been treated with the immunosuppressant cyclosporine A (180).
It is obvious that novel, disease mechanism targeted treatments are needed, and these should probably be systemic in order to target multiple organ dysfunctions. Moreover, perhaps these treatments should be commenced early in life in order to prevent, rather than just treat, the disease. As discussed earlier, impaired SERCA is at the core of DD. Pharmacological activation of SERCA may therefore ameliorate DD-related symptoms. A recent study (36) showed that CDN1163, a SERCA activator that directly binds to the SERCA2b enzyme to activate its Ca2+-ATPase activity probably via an allosteric mechanism (181), improved metabolic parameters in vivo in a type 2 diabetes mouse model (ob/ob) (36). Moreover, Savignac et al. (36) showed that ER stress and associated impairment of cell-to-cell adhesion in patients with DD-derived primary keratinocytes were rescued by the pharmacological chaperone miglustat, which is a clinically available drug. Thus, candidates that affect ER calcium or ER stress exist and could be included in clinical trials.
Since DD is usually inherited, a method of preventing the disease rather than just treating it is to perform preimplantation genetic diagnosis of fertilized embryos, in which 1 or 2 cells (blastomeres) are removed from the preimplantation embryo at the 6–10-cell stage (day 3 of development), thus allowing replacement into the uterus of unaffected embryos. This is, of course, a difficult ethical decision; however, as the systemic aspects of DD become more established more parents are likely to consider this alternative and have the right to be fully informed about it by their dermatologists and referred for genetic counselling (17). In our experience, most patients are unaware of this option. Moreover, it is important to perform genetic testing on all patients, as some do not have pathological mutations and, in our experience, some patients choose not to have children in order not to transmit the disease. While more studies are needed on the systemic aspects of DD, patients should be fully informed on the systemic aspects of their disease, although we cannot currently offer any treatments that target organs other than the skin.
While research on the systemic aspects of DD is still in its infancy, it is becoming clear that DD is a multi-organ systemic condition (Fig. 3). This not surprising, considering the overwhelming experimental evidence of the importance of SERCA2 in physiology and pathophysiology. While more data is needed, treating physicians should be aware of the risk of extracutaneous manifestations. Future research would benefit from more registry studies, as well as from systemic large-scale DD cohort studies, in order to examine associations as well as to find direct experimental evidence of other organ dysfunctions and diseases.
Fig. 3. Overview of emerging systemic extracutaneous involvement in Darier disease.
The authors would like to extend sincere thanks and appreciation to the participating patient in Fig. 1. The patients in this manuscript have given written informed consent to publication. The authors are also grateful to the funding agencies (Vetenskapsrådet, Hudfonden, Svenska Sällskapet för medicinsk forskning, ALF medicin Stockholm, Jeanssons stiftelse, Tore Nilssons Stiftelse) for their support. Jakob Wikström was supported by a Wallenberg clinical fellow grant from the Marianne and Marcus Wallenberg foundation.
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize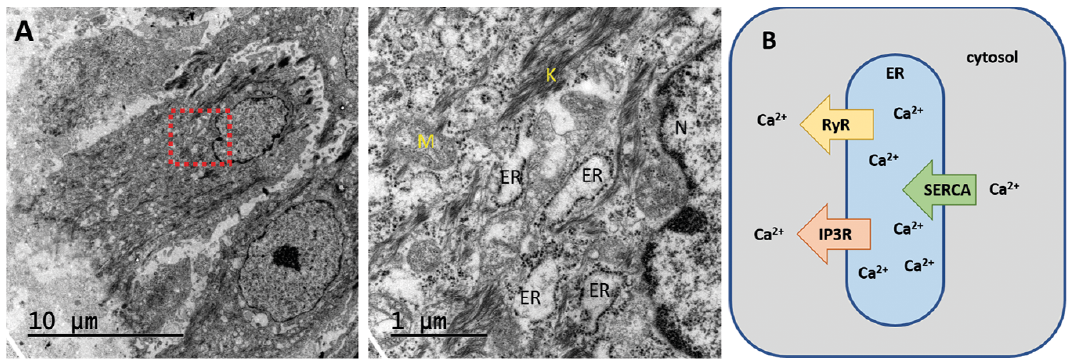 Click to show fullsize
Click to show fullsize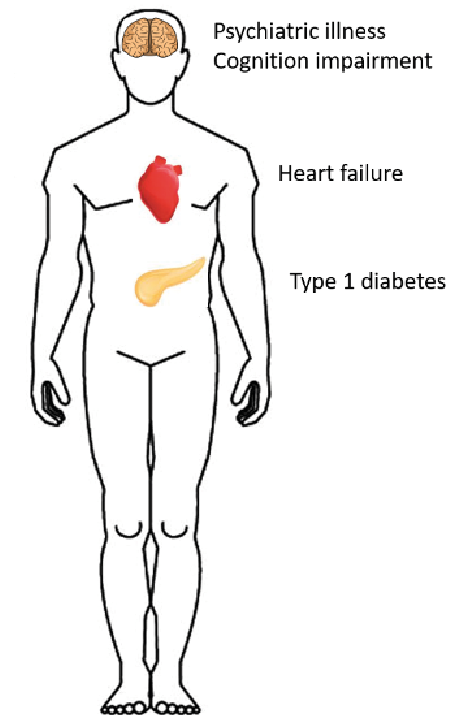 Click to show fullsize
Click to show fullsize