


1INSERM U976 Saint Louis Hospital Paris, 2INSERM U981, Gustave Roussy Cancer Campus, Paris Saclay University, Villejuif, 3Paris University, Department of Pathology, 4Department of Tumor Genomics and Pharmacology, DMU BioGem, Saint Louis Hospital, Paris, 5Paris University, Department of Dermatology, Saint Louis Hospital, FR-75010 Paris, France. *E-mail: nicole.basset-seguin@aphp.fr
Accepted Apr 1, 2021; Epub ahead of print Apr 13, 2021
Acta Derm Venereol 2021; 101: adv00434.
doi: 10.2340/00015555-3797
Gorlin syndrome comprises multiple basal cell carcinomas (BCCs), basal cell naevi (BCNs), odontogenic keratocysts, skeletal abnormalities and other features (1).
Identification of the PTCH1 gene as the candidate gene for Gorlin syndrome demonstrated that activation of the Hedgehog (Hh) pathway is the key driver in the physiology and pathology of BCC formation (2). Gorlin syndrome has been linked to germline mutations of various members of the Hh pathway, including mainly PTCH1 and also PTCH2 and SUFU; however, to date, SMO germline mutations have not been described. Recently, a case of Gorlin syndrome type 1 mosaicism presented a somatic mutation of SMO L412F (3). This mutation has been described as oncogenic, conferring constitutive activation of the Hh pathway (4).
This paper describes a second case of Gorlin syndrome mosaicism with the same SMO L412F mutation observed in affected skin. A detailed molecular assessment of multiple BCNs and BCCs was performed to analyse the oncogenic steps of BCC formation in this patient.
A 33-year-old woman has been followed for multiple BCCs since the age of 27 years. There was no family history of BCC.
Physical examination revealed hypertelorism and frontal bossing. Odontogenic cysts, cerebral calcification and palmoplantar pits were absent. The right side of her face and neck was covered with multiple skin-coloured papules, which, on excision and haematoxylin and eosin (H&E) staining, showed either BCC (Fig. 1A and C) or BCN (Fig. 1B and D). The patient provided informed signed consent to perform molecular analysis of the frozen and formalin-fixed paraffin-embedded (FFPE) tumour samples. BCCs presented large tumours of basaloid cells invading the dermis of the nodular, micronodular or infundibulocystic subtypes, with mitotic activity and slight cytological atypia (Fig. 1C). BCN consisted of tiny arborescent buddings of cytologically bland basaloid cells attached to the epidermis or replacing the hair follicles (Fig. 1D).
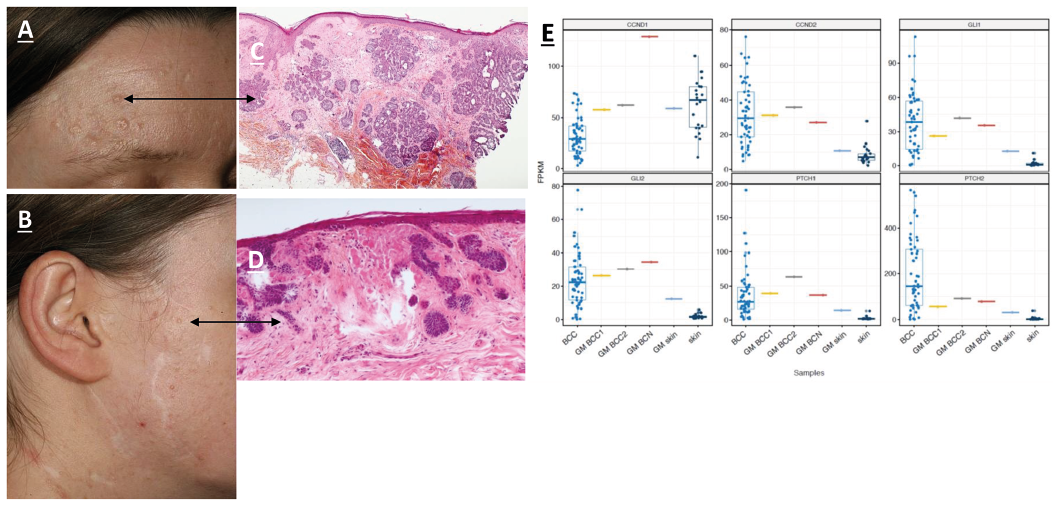
Fig. 1. Clinical and histological presentation of basal cell naevi (BCN) and basal cell carcinoma (BCC). (A) Nodular BCC in the vicinity of other BCN and BCC; (B) facial skin coloured papule; (C) haematoxylin and eosin (H&E) staining showing a micronodular BCC; (D) H&E staining showing BCN; (E) expression of cyclins and major BCC markers in the Gorlin mosaic (GM) patient and sporadic BCCs. BCC: sporadic BCCs from Bonilla et al. 2016; GM BCC1/2: BCC from the GM patient; GM BCN: BCN from the GM patient; GM_skin: normal skin from the GM patient; skin: normal skin samples from Bonilla et al. (6). Fragments per kilobase per million (FPKM), relative level of expression measured as FPKM.
Next-generation sequencing (NGS) was performed on 11 BCCs and 7 formalin-fixed paraffin-embedded (FFPE) BCNs. H&E staining of the biopsy samples, except for 2 lesions that required microdissection, revealed a tumour cellularity range from 30% to 50% for BCC. DNA was extracted as described previously (5) using a Maxwell RSC DNA FFPE kit (Promega, Charbonnières-les-Bains, France) according to the manufacturer’s instructions.
A custom-designed AmpliSeq™ NGS panel (Thermo Fisher Scientific, Les Ulis, France) of 95 genes (Appendix S1), including genes described in all skin carcinomas (6–8), was used for comprehensive genotyping of the tissue biopsies and cutaneous control samples (normal skin of the patient), as described previously (5).
NGS analysis revealed that all tumours harboured a common SMO p.L412F mutation. Additional mutations, including PTCH1 and MYCN, were observed in BCN and, more frequently, in BCC. All mutations harboured an ultraviolet (UV) signature (e.g. C>T transitions). No mutations were detected in adjacent or distant normal skin or in the blood.
Four frozen tissue samples from the patient (1 BCN, 2 BCC and 1 normal skin) were studied by RNA-seq. RNA was extracted using an AllPrep RNA kit (Qiagen, Valencia, CA, USA) according to the manufacturer’s protocol. Extracted RNA samples were sequenced using a BGISEQ-500 instrument (BGI, Shenzhen, China) and processed by a standard RNA-seq pipeline that included read alignment by STAR (9) and read count quantification by HTSeq (10) with subsequent conversion to fragments per kilobase per million (FPKM). Data published previously by Bonilla et al. (6), representing BCC samples (n = 57) and normal skin samples (n = 22), were used to compare the levels of expression of a panel of major BCC markers and cyclins in the patient samples.
A strikingly elevated level of CCND1 was found in a BCN sample, which had a level of CCND2 comparable to that found in other tumours. The level of Hh activation, measured as the expression of major BCC markers (GLI1, GLI2, PTCH1 and PTCH2), was similar in BCN and BCC from the current patient and was within the range of values reported previously for BCC samples (Table SI).
Segmental Gorlin syndrome is rare and corresponds to a type 1 mosaicism resulting from a postzygotic mutation. A first case of segmental Gorlin syndrome involving an activating (p.L412F) SMO mutation was reported recently (3). The mutations in PTCH1, PTCH2 or SUFU, which are the main germline mutations in classical Gorlin syndrome, were not detected in the blood or skin of the patient. We report here the second case of segmental Gorlin syndrome, presenting the same activating SMO mutation only in affected skin. This case adds to the list of mosaic hedgehog spectra implicating the SMO gene (11). Interestingly, the same SMO mutation was observed in affected organs with various phenotypes, including Happle–Tinschert syndrome (11), Curry-Jones syndrome and segmental Gorlin syndrome. These syndromes have many common clinical manifestations and probably represent differential expression of the same disease as they share an identical genetic trait. Phenotypic variability often detected in genetic mosaicisms was reported to depend on the mutation load and involved tissues (12).
In the current patient, the SMO p.L412F mutation was present in all tumour types, but not in distant or adjacent normal skin, which is similar to the pattern observed in the first reported case. This SMO p.L412F mutation was reported only in segmental cases and was never observed in constitutive Gorlin syndrome, suggesting that the germline mutation is lethal. The p.L412F mutation was the only event detected in some tumours, both BCN and BCC, suggesting that it is sufficient for tumour formation within the tested gene panel. The number of mutations did not significantly differ between BCN and BCC, although additional mutations were more frequent in BCC (7/11; 64%) than in BCN (1/7; 14%).
The SMO p.L412F has been reported as an oncogenic driver (4) associated with resistance to vismodegib in sporadic BCC, and tumour regrowth was associated with PTCH1 levels in the context of this oncogenic mutation, as partial or total PTCH1 loss affected drug sensitivity, as suggested in the study of Sharpe et al. (13). Thus, the current patient was not treated with hedgehog inhibitors. However, vismodegib was used in the first reported case, and the treatment appeared to be effective. In several tumours, the somatic variant of SMO co-occurred with PTCH1 variants. This phenomenon was reported previously by Bonilla et al. (6) in sporadic BCC and may suggest an additive effect of these mutations manifested as a reduced interaction between PTCH1 and SMO.
At the RNA level, activation of the Hh pathway was observed in both BCC and BCN and was comparable to that observed in sporadic BCC. Unlike an SMOM2 mouse model, which manifested elevated expression of GLI and cyclins D1 and D2 in BCC and not in BCN, we did not observe a similar difference in our samples (14). In contrast, the results for the current patient at the RNA level showed that the highest levels were detected in a single BCN compared with that in the 2 BCC samples tested; however, this result requires careful interpretation because only a single BCN was analysed.
This case report confirms that mosaic (post-zygotic) activating SMO mutations contribute to Gorlin syndrome/BCNS. This mutation appears to be sufficient for tumour formation in both BCN and BCC. Comparison of BCN and BCC at the genetic and transcriptional levels indicates that these tumours have similar molecular alterations, suggesting that they are part of the same tumour process and differ only in tumour size and mutation load.
The authors thank Genentech/Roche for their financial support and the patient for her cooperation.
SN was supported by grant Foundation ARC 2017, Foundation Gustave Roussy and Swiss Cancer League KFC-3985-08-2016
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize