


1Department of Dermatology, College of Medicine, Hallym University, Kangnam Sacred Heart Hospital, Seoul, 2College of Pharmacy, Gachon University, Incheon and 3Department of Anesthesiology and Pain Medicine, College of Medicine, Hallym University, Hangang Sacred Heart Hospital, Seoul, South Korea
#These authors contributed equally to this work.
Carvacrol, a natural transient receptor potential vanilloid-3 activator, has been reported to cause pruritus in mice. This study aimed to evaluate the effects of carvacrol and various antipruritic agents in humans. A stimulation test with carvacrol, β-alanine, and histamine was performed. After application of the pruritic solutions, the skin was stimulated with pinpricks. In inhibition test A, Forsythia suspensa extract, containing forsythoside B (a transient receptor potential vanilloid-3 inhibitor), was applied by pricking prior to stimulation with pruritogens. In inhibition test B, olopatadine solution, tacrolimus ointment, and Scutellaria baicalensis root extract were applied, and carvacrol was applied to the same region. Carvacrol induces moderate pruritus in humans. The pruritus was relieved by Forsythia suspensa extract and olopatadine solution after 20 min of application and by tacrolimus ointment and Scutellaria baicalenis extract after 24 h of application. These results suggest that carvacrol is a pruritogen in humans, and that carvacrol-induced pruritus is inhibited by various antipruritic agents.
Key words: itch; pruritus; carvacrol; transient receptor potential vanilloid-3; thymic stromal lymphopoietin; oregano.
Accepted Jun 28, 2021; Epub ahead of print Jun 29, 2021
Acta Derm Venereol 2021; 101: adv00517.
doi: 10.2340/00015555-3855
Corr: Hye One Kim, Department of Dermatology, College of Medicine, Hallym University, Kangnam Sacred Heart Hospital, Seoul, South Korea. E-mail: hyeonekim@gmail.com
Pruritus is a distressing sensation that triggers a desire to scratch and has a significant impact on quality of life. Current evidence strongly suggests that transient receptor potential ion channels in human skin play a key role in many types of pruritus. This study examined the putative effect of transient receptor potential vanilloid-3 channel activation on pruritus. The effect was measured by comparing the subjective degree of pruritus with that of other pruritogens. The results suggest that transient receptor potential vanilloid-3 activation is a potential pruritogen, and novel treatment modalities can be suggested by inhibiting this channel.
Pruritus is defined as an unpleasant sensation that elicits a desire to scratch (1). It is known to impair quality of life and is a major symptom of chronic inflammatory diseases, such as atopic dermatitis (2). Histamine is one of the most well-known pruritogens, and antihistamines can be used to manage pruritus. Studies on the pathophysiology of pruritus have discovered various pruritogens other than histamine, and antagonism of these pruritogens could be developed to provide new treatments for pruritus.
The transient receptor potential (TRP) channels are a group of non-selective cation channels, which are mostly expressed on the plasma membrane of cells. Although other TRP channels are normally expressed in sensory neurones, a specific subtype called transient receptor potential vanilloid-3 (TRPV3) is abundantly expressed in keratinocytes (3). Activation of TRPV3 in keratinocytes regulates the proliferation, differentiation, and apoptosis of human skin. TRPV3 channels can be activated both by warm temperatures, and by common materials. For example, well-known chemical TRPV3 agonists are carvacrol (a plant-derived compound from oregano) and 2-aminoethoxydiphenyl borate, a synthetic agonist (4).
The association between TRPV3 activation and pruritus is typically observed in Olmsted syndrome. The syndrome is characterized by multiple gain-of-function mutations in the TRPV3 gene. As the TRPV3 channel is involved in keratinocyte migration and wound-healing via nitric oxide, the syndrome is also characterized by hyperkeratosis, keratoma, and diffuse alopecia (5–7). In addition, Olmsted syndrome is accompanied by extreme pruritus with immune dysfunction, such as hyper-IgE and chronic eosinophilia (8). In addition to Olmsted syndrome, other pruritic dermatological disorders show an association with TRPV3. Indeed, overactive TRPV3 has been reported in the pathogenesis of atopic dermatitis in mice (9). In our previous studies, hypertrophic scars with post-burn pruritus showed increased expression and activity of TRPV3 in keratinocytes, and the activation of TRPV3 induced Ca2+ influx in cultured keratinocytes derived from pruritic burn scars (10, 11). Activation of TRPV3 channels in human primary keratinocytes and human primary dermal fibroblasts induces transcription of thymic stromal lymphopoietin (TSLP), a cytokine that can initiate various allergic diseases (10–12). Known inhibitors of the TRPV3-TSLP pathway include forsythoside B (13), tacrolimus (14) and baicalein (15). Forsythoside B selectively inhibits the TRPV3 channel, putatively by blocking ion passage; tacrolimus prevents the dephosphorylation of nuclear factor of activated T-cells (NFAT); and baicalein can block the TSLP signalling pathways.
An animal model study was conducted to determine whether TRPV3 activation causes pruritus in vivo (16). The study found that intradermal injection of carvacrol in mice induced scratching in a dose-dependent manner. Another in vivo study provided further evidence for the effect of forsythoside B on attenuating the pruritus caused by carvacrol in mice (13). This study suggests that specific inhibition of TRPV3 by forsythoside B can be a potential treatment for alleviating pruritus. However, it is not yet certain whether TRPV3 activation leads directly to pruritus in humans. Therefore, to answer this question, the aims of this study were to determine whether a TRPV3 agonist (carvacrol) can induce pruritus in humans, and whether inhibitors of TRPV3 and TSLP can attenuate carvacrol-induced pruritus in humans.
Reagents
Carvacrol 98% and β-alanine 99% were purchased from Sigma-Aldrich (St Louis, MO, USA). Carvacrol (49%) was prepared by mixing 0.5 mL ethanol with 98% carvacrol at a volume of 0.5 ml in a 1 ml syringe. β-alanine (50%) was prepared by dissolving 100 g β-alanine 99% crystal in 99 g normal saline. Histamine was obtained from Tokyo Chemical Industry (Tokyo, Japan). Pazeo® (olopatadine hydrochloride ophthalmic solution, 0.7%, Novartis), Protopic® (tacrolimus 0.1% ointment, Astellas Pharm Inc., Tokyo, Japan), Forsythia suspensa extract (33%, Hawaii Pharm, Honolulu, USA), and Scutellaria baicalensis root extract (33%, Herbal Terra, Hawaii, USA) were used as inhibitors of pruritus. These extracts were chosen because they are a source of specific TRPV3 pathway inhibitors. Scutellaria baicalensis root extract is a source of baicalein, a TSLP pathway inhibitor, and Forsythia suspensa extract contains the specific TRPV3 inhibitor forsythoside B. Tacrolimus ointment, another inhibitor of the TRPV3-NFAT-TSLP pathway, was also included in the study, as well as the antihistamine olopatadine. This latter was to determine whether the TRPV3 pathway has any association with histamine.
Methods
The study was approved by the Institutional Review Board (IRB) of Hallym University Kangnam Sacred Heart Hospital, Seoul (IRB number 2019-05-018).
Fifteen healthy Asian subjects over 19 years of age were recruited for this study. Subjects with pruritus, medication use, and current allergic or dermatological disorders were excluded. Written informed consent was obtained from all participants prior to their participation in the study. The mean age of the subjects was 29.07 years (range 26–39 years) and the ratio of males to females was 7:8. The study was conducted in 3 parts: a stimulation test and inhibition tests A and B, all of which were conducted in a double-blind manner. To facilitate this, the experiment and the numerical rating scale (NRS) recordings were conducted by different people. In the stimulation test, the provocations were performed with carvacrol (TRPV3 activator, 49% and 98% weight by volume in ethanol), β-alanine (50%), histamine (1%), and ethanol (10%; control) by skin-prick test. Five areas were marked, and applications were carried out on the volar surfaces of the forearms. A 3-cm distance between the 2 trigger locations was set in order to minimize false-positive reactions. One drop of each pruritic solution was applied to the skin and immediately stimulated with pin-pricks. This method allowed the pruritogens to penetrate through the epidermal barrier. The pruritus intensity was evaluated in a continuous manner using the NRS, ranging from 0 (no pruritus) to 10 (worst pruritus). Subjects rated pruritus intensity for 10 min at 1-min intervals after the application of pruritogens. Before and after the study, the erythema index (EI) was measured using a spectrophotometer® (Cortex Technology, Hadsund, Denmark). The measurements were conducted in a room at constant temperature (20–24°C) and humidity (28–38%). The presence of erythema, oedema, or other skin lesions was evaluated by an investigator.
In inhibition test A, TRPV3 inhibitor (forsythia extract) was applied precisely to the same regions provoked by the skin prick test. After 30 min of application of inhibitor, the same substances used for the stimulation test were applied to the same regions. The NRS for pruritus was evaluated in the same manner as the stimulation test.
Inhibition test B determined the effect of inhibitors in reducing pruritus induced by carvacrol 98%. Scutellaria baicalensis extract, tacrolimus ointment, olopatadine solution, and forsythia extract were loaded onto IQ UltimateTM 1×1 cm chamber sheets (Chemotechnique Diagnostics, Vellinge, Sweden) and subsequently applied over the skin. Twenty minutes and 24 h after application of inhibitor, carvacrol 98% solution was applied to the same regions. The NRS score for pruritus was evaluated in the same manner. To avoid interference inhibition tests A and B were conducted on different days.
Statistical analyses
Continuous data are expressed as means ± standard deviations. Student’s t-test for continuous variables was used to determine the significance of differences. Repeated measures analysis of variance (ANOVA) was used to analyse the changes in itch intensity with respect to time. The statistical significance of any differences between groups was determined using ANOVA with Bonferroni post-hoc analysis. The significance level for all analyses was set at p < 0.05. All statistical analyses were conducted using PASW Statistics 18 (SPSS Inc., Chicago, IL, USA).
Pruritus induced by carvacrol in a dose-dependent manner
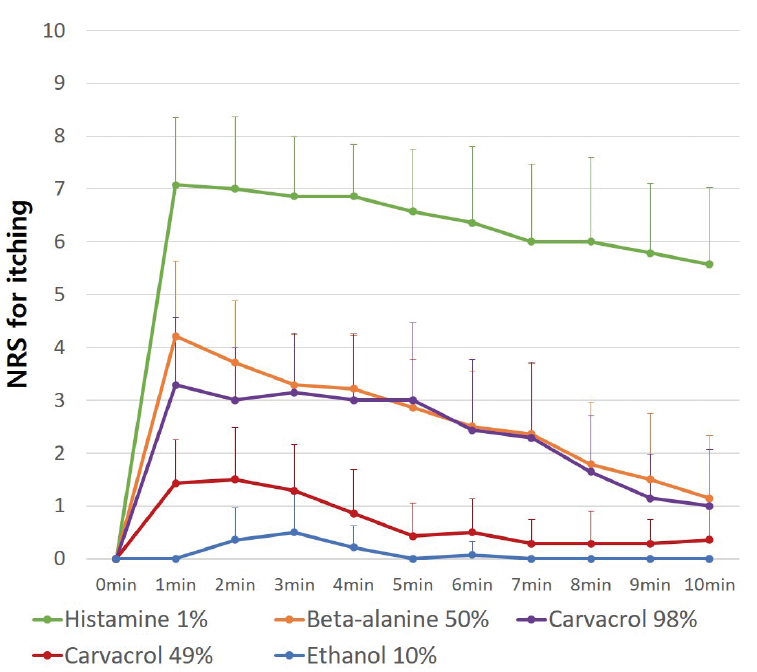
Of the pruritogens studied, histamine induced the most severe pruritus (Fig. 1). ß-alanine and carvacrol induced moderate pruritus, which were not significantly different from each other. Carvacrol induced pruritus in a dose-dependent manner. The pruritus decreased gradually over time after stimulation by pruritogens, and the changes in pruritus with respect to time were significantly different (p = 0.000). All pruritogens significantly induced pruritus compared with the vehicle (ethanol) (Fig. 1 and Fig. 2, p < 0.001). This was confirmed by the mean itch intensity. The mean itch intensity caused by carvacrol 98% was higher than that caused by carvacrol 49% (2.39 ± 0.95 vs 0.72 ± 0.49, Fig. 2, p = 0.000). Stimulation by pruritogens other than histamine did not increase the erythema index (EI) or cause wheal-and-flare reactions. Visually, erythema, oedema, or other skin lesions were not present, except for at the histamine stimulation site.
Fig. 1. Trend line of numerical rating scale (NRS) for pruritus induced by several pruritogens in stimulation test. Histamine induced the most severe pruritus, followed by ß-alanine and carvacrol. Carvacrol caused pruritus in a dose-dependent manner. The changes in pruritus with respect to time were significantly different (p = 0.000). The pruritus decreased over time after stimulation with pruritogens. *p-values were determined using repeated measures analysis of variance (ANOVA). Analysis included 15 healthy Asian subjects.
Fig. 2. Mean itch intensity caused by several pruritogens in stimulation test. ###All pruritogens induced pruritus significantly more than the control (ethanol), p < 0.001. ***p < 0.001, NRS: numerical rating scale. *p-values were determined using the Student’s t-test. Analysis included 15 healthy Asian subjects. The integration time for the mean NRS rating was 10 min.
Effect of forsythia extract on the reduction of pruritus owing to histamine and carvacrol
Application of forsythia extract prior to stimulation reduced pruritus intensity (Fig. 3A–D). The effect of forsythia extract was statistically significant for histamine 1% solution-induced pruritus (6.41 ± 1.19 vs 5.08 ± 0.83, Fig. 3A, p = 0.015) and carvacrol 98% solution-induced pruritus (0.976 g/ml) (2.39 ± 0.94 vs 0.97 ± 0.73, Fig. 3C, p = 0.001). Both pruritogens acted in a dose-dependent manner, but the forsythia extract (50% solution) was significantly more effective in reducing the pruritus caused by carvacrol 98% solution (2.39 ± 0.94 vs 1.47 ± 1.06, Fig. 3C, p = 0.039). In contrast, the effects of forsythia extract did not significantly reduce pruritus caused by ß-alanine (p = 0.523) or carvacrol 50% solution (p = 0.200).
Fig. 3. Mean itch intensity induced by (a) histamine 1%, (b) β-alanine 50%, (c) carvacrol 98% and (d) carvacrol 49% after application of forsythia extract solution for 30 min. The pruritus induced by histamine 1% and carvacrol 98% solution was relieved by application of forsythia extract 33% solution. *p < 0.05, **p < 0.005, ***p < 0.001. *p-values were determined using 1-way analysis of variance (ANOVA) with Bonferroni post-hoc analysis. F-value: 6.383 (a: histamine), 6.304 (b: β-alanine), 12.472 (c: carvacrol 98%), 3.109 (d: carvacrol 49%). Analysis included 15 healthy Asian subjects over 19 years of age. The integration time for the mean numerical rating scale (NRS) rating was 10 min.
Effect of antipruritic agents on the reduction of pruritus due to carvacrol
Olopatadine solution and forsythoside B significantly reduced carvacrol-induced pruritus, whereas other antipruritic agents did not (Figs 4 and 5). The trend line was similar to that of the stimulation test, as pruritus gradually decreased over time. The inhibitory effect of olopatadine solution (2.64 ± 0.77 vs 1.26 ± 0.92, p = 0.000) and forsythoside B (2.64 ± 0.77 vs 0.94 ± 0.61, p = 0.001) supported the inhibitory effect of antagonisms on histamine and TRPV3 calculated by the mean itch intensities (Fig. 5). Meanwhile, the Scutellaria baicalensis root extract (p=0.867) and tacrolimus ointment (p = 0.480) did not immediately reduce carvacrol-induced pruritus. One day after application of inhibitor, the tacrolimus ointment (2.63 ± 0.77 vs 1.41 ± 0.53, p = 0.000) and Scutellaria baicalenis root extract (2.63 ± 0.77 vs 1.22 ± 0.29, p = 0.000) reduced the pruritus due to carvacrol.
Fig. 4. Trend line of numerical rating scale (NRS) for pruritus induced by carvacrol with pre-application of inhibitors for 20 min in inhibition test B. The inhibitors were loaded onto the patch chamber and applied before carvacrol stimulation. Olopatadine solution and forsythia extract significantly reduced the carvacrol-induced pruritus. The trend line is similar to stimulation test showing gradual decrease with time. Analysis included 15 healthy volunteers.
Fig. 5. Mean itch intensity induced by carvacrol 98% solution after application of several inhibitors for 20 min (a) and 24 h (b). The inhibitors were loaded onto the patch chamber and applied before carvacrol stimulation. The pruritus of carvacrol 98% solution was relieved by the application of olopatadine solution and forsythia extract 98% solution after 20 min and by Scutellaria baicalenis extract and tacrolimus ointment after 24 h. ***p < 0.001. *p-values were determined using Student’s t-test. Analysis included 15 healthy Asian subjects. The integration time for the mean numerical rating scale (NRS) rating was 10 min.
The pruritogenic effect of carvacrol has been reported previously in a mouse model (16). Specifically, intradermal injection of carvacrol-induced scratching behaviours in mice in a concentration-dependent manner, and the episodes of scratching induced by carvacrol were significantly fewer in number in TRPV3-knockout mice. Since carvacrol can activate both TRPV3 and TRPA1, this suggests that pruritus was not completely abolished in TRPV3-knockout mice. However, the scratching time was reduced predominantly in the sustained phase, in which TRPV3 mainly acts. Another study in a mouse model reported that forsythoside B can significantly attenuate pruritus induced by either carvacrol or histamine, via inhibition of TRPV3 (13). Indeed, forsythoside B showed dose-dependent inhibition of TRPV3, resulting in a reduction in scratching episodes in mice (13). In addition, our previous study showed that carvacrol, which activates both TRPA1 and TRPV3, induces pruritus in humans with burn scars (17).
The aim of this study was to determine whether pruritus is triggered by the application of carvacrol, compared with other pruritogens, in humans. Since the effect of the TRPV3 activator on human skin has not been evaluated previously, the current study aimed to select certified cosmetic ingredients as pruritogens. Carvacrol and forsythia extracts containing forsythoside B were selected, which are both of natural origin and have been used as cosmetic ingredients. Baicalein, a major component of Scutellaria baicalensis, has been reported to block TSLP signalling pathways (18). Scutellaria baicalenis root extract was selected as an inhibitor because it contains baicalein as a major component, and its safety as a cosmetic ingredient has been proven previously. In addition, the skin prick test was chosen instead of intradermal injection for minimal systemic absorption. Consistent with a previous animal model study (16), the experiment was started with low concentrations (0.01%, 0.1% and 1%) of carvacrol. However, the lower concentration did not provoke pruritus when the skin prick method was used; therefore, we adjusted the concentration continuously and chose a more appropriate concentration (49% and 98%, respectively).
The intensity of pruritus elicited as a result of carvacrol was similar to that of ß-alanine and was concentration-dependent. The current study results are similar to those of another mouse model study, which concluded that carvacrol acts as a pruritogen by activating TRPV3. The current study assessed the intensity of carvacrol-induced pruritus on human skin compared with pruritus induced by other pruritogens. The forsythia extract significantly attenuated pruritus caused by the histamine and carvacrol 98% solutions. Meanwhile, carvacrol-induced pruritus was reduced by olopatadine and forsythia extracts after 20 min of pre-application. The finding that forsythoside B can be an effective treatment for managing pruritus was consistent with previous results. Furthermore, the current study attempted to find inhibitors to reduce carvacrol-induced pruritus over time. Olopatadine and forsythia extracts were effective by 20 min after application, whereas the tacrolimus ointment and Scutellaria baicalenis extract did not show any effect 20 min after application. The inhibitory effects of tacrolimus ointment and S. baicalenis extract were observed 24 h after application. This is presumed to be because tacrolimus, a calcineurin inhibitor, takes several hours or more to inhibit the transcription of TSLP by inhibiting the nuclear factor of activated T-cells (NFAT) pathway. Calcium influx into keratinocytes triggers calcineurin-dependent dephosphorylation and nuclear translocation of NFAT, which induces transcription of TSLP (15). Meanwhile, the inhibitory effects of olopatadine and forsythia extracts disappeared 24 h after application. It is assumed that the direct inhibition of TRPV3 channels by forsythia extract could not persist for 24 h, and the olopatadine solution was not effective in inhibiting direct histamine release for 24 h.
TRP channels are ion channels that play an important role in pruritus (19). Pruritus associated with TRPV3 activation has previously been observed in various dermatological diseases. Olmsted syndrome, a syndrome characterized by gain-of-function mutations in TRPV3, induces extreme pruritus with elevated immunoglobulin E (IgE) levels and chronic eosinophilia. Atopic dermatitis and post-burn pruritus also show overactive TRPV3 expression (9–11). However, pruritus induction experiments have not been conducted in healthy people. This study evaluated the effect of carvacrol on pruritus by measuring its intensity and determining the materials that can attenuate pruritus. This may help elucidate the mechanism by which pruritus can be transmitted from keratinocytes to neurones.
The hypothesized mechanism by which pruritus is transmitted from keratinocytes to sensory neurones is shown in Fig. 6 (17). Initially, mast cells release tryptase, which binds to protease activated receptor 2 (PAR2) in keratinocytes. Activation of PAR2 leads to the downstream stimulation of multiple signalling pathways (protein kinase A [PKA], PKB, PKC). These signals further trigger the activation of TRPV3, which induces continuous calcium influx into the keratinocyte. The increased calcium concentration with the cell augments activation of the calcium-calmodulin pathway, resulting in dephosphorylation of NFAT, which then leads to increased TSLP expression. Once TSLP is released from the keratinocytes, the nearby sensory nerve fibres with cognate TSLP receptors bind to TSLP, which results in generation of an electrical signal. The excitation of the sensory dorsal root ganglion is relayed into the dorsal horn of the spinal cord, and finally to the brain. The association between TSLP release by TRPV3 activation and post-burn pruritus was established in our previous study (20).
Fig. 6. Hypothesized mechanism of transient receptor potential vanilloid-3 (TRPV3) associated pruritus. Figure modified from graphic abstract in reference (20). *(number) refers to the article number used as a reference in this study. PAR2: protease-activated receptor 2; PKA: protein kinase A; PKC: protein kinase C; PKD: protein kinase D; TRPV3: Transient receptor potential vanilloid-3; NFAT: nuclear factor of activated T cells; DRG: dorsal root ganglion; TSLP: thymic stromal lymphopoietin; H1R: histamine 1 receptor; TRPA1: transient receptor potential ankyrin 1.
The current study revealed that TRPV3 activation by carvacrol induced moderate pruritus, and TRPV3 inhibition by forsythia extract reduced the pruritus induced by carvacrol. In addition, histamine can be a potential mediator in pathways, since the pruritus caused by carvacrol was reduced by olopatadine solution, and pruritus caused by histamine was reduced by forsythia extract. In a previous mouse model study (21), hairless mutants (DS-Nh) have been reported to express TRPV3 gain of function mutations in keratinocytes. In this previous study, it was found that histamine levels in the skin of control mice (DS) were lower than those in the skin of hairless mutants (DS-Nh) of the same age. Although the precise mechanisms have not yet been revealed, the ability of keratinocytes to release histamine directly was observed in another study (22, 23). This study showed that intradermal injection of α-melanocyte-stimulating hormone (α-MSH) induces itch in mast cell-deficient mice. In addition, the application of α-MSH also induced the release of histamine from both mast cells and keratinocytes. Since the activation of melanocortin receptors by α-MSH increased the intracellular calcium levels, we also suggested that intracellular calcium levels may contribute to the direct production of both histamine and TRPV3 in keratinocytes.
In addition, this study determined the inhibitory effect of tacrolimus ointment on carvacrol-induced pruritus. Tacrolimus prevents the dephosphorylation of NFAT by binding to the FK-binding protein in calcium-calmodulin signalling (14), and it is this inhibitory effect on dephosphorylation that attenuates carvacrol-induced pruritus. Since our previous study (10) has revealed that TRPV3 activation stimulates Ca2+ influx in vivo, the inhibitory effect of tacrolimus ointment is evidence that the NFAT signalling pathway is involved in this pathway. The inhibitory effect of Scutellaria baicalensis extract may be due to the inhibition of the TSLP signalling pathway.
Carvacrol is known to both activate TRPV3 and inhibit TRPM7 (24). Although TRPM7 is ubiquitously expressed among tissues and cells, it is not specifically expressed in keratinocytes. This suggests that the effect of TRPV3 is more dominant in keratinocytes than that of TRPM7. Forsythoside B has been reported to inhibit inflammatory cytokines, such as tumour necrosis factor alpha (TNF-α), interleukin (IL)-6 and nuclear factor (NF)-κB (25). However, the inhibitory effect of forsythoside B on these cytokines was confirmed 24 h after pretreatment with forsythoside B. Since the current study evaluated the inhibitory effect of forsythoside B for 10 min after pruritogen stimulation, the effect of other inflammatory cytokines was minimized. Baicalein has also been reported to have inhibitory effects on other enzymes, including beta-secretase 1 (BACE1) and acetylcholinesterase (AChE), in addition to TSLP (18, 26). This effect can distort the experimental results by hampering the TSLP pathway through unintended interaction with other metabolic pathways. However, since BACE1 is usually distributed in nerve cells, involving the formation of myelin sheaths (27), and AChE is usually found at postsynaptic neuromuscular junctions for terminating neuronal transmission, these inhibitory effects would be smaller than the effect on TSLP. Additional studies correcting these variables would be helpful in proving the effect.
Study limitations
Remarkably, carvacrol has been proven to be a pruritogen, along with other inhibitors, on human skin. However, this study has several limitations. First, the sample size was small. Secondly, the study had to use delivery systems or materials whose safety has been proven. Although an intradermal test or lower concentration of solution might be more accurate, we could not use them due to consideration of participants’ safety. Additional experiments using several lower concentration solutions in intradermal tests would help to determine the results in detail. Thirdly, since the NRS is a subjective evaluation, it could be influenced by other variables. To address this limitation, this experiment was performed with a double-blinded and vehicle-controlled design, checked the NRS at an accurate time, and excluded participants who had a recent history of pruritus.
Conclusion
This study investigated the effect of carvacrol (a TRPV3 activator) and forsythia extract (a TRPV3 inhibitor) on pruritus. The results suggest that carvacrol, a natural TRPV3 activator, is a pruritogen in humans as well as in mouse models. Since the effect of inhibitors in reducing the pruritus induced by carvacrol was assessed, we can consider the mechanism of TRPV3-associated pruritus.
This study was supported by grants from the National Research Foundation of Korea (NRF-2017R1A2B4006252 and NRF-2018R1C1B6007998), by the Korea Centers for Disease Control and Prevention (KCDC 2020-ER6714-00), and by the Korea Healthcare Technology R&D Project (HI17C0597) funded by the Ministry of Health & Welfare, Republic of Korea, by the Hallym University Research Fund.
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize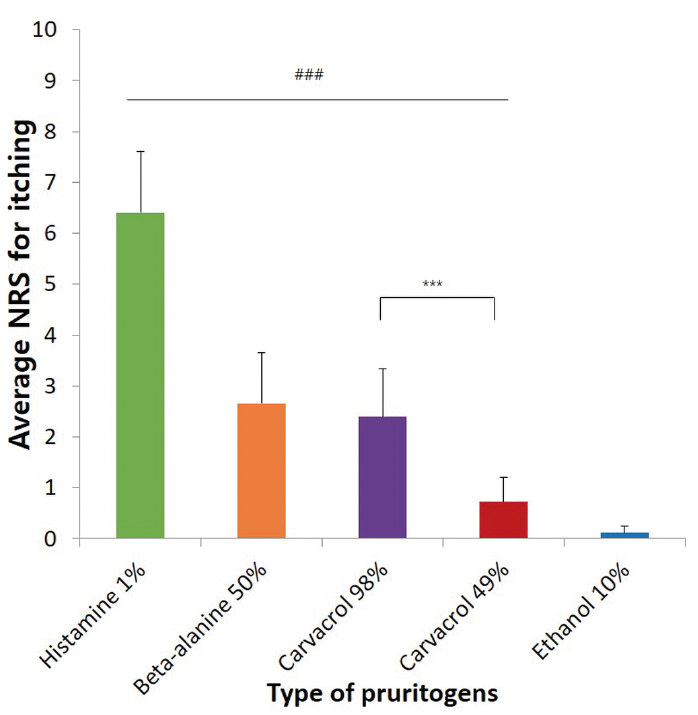 Click to show fullsize
Click to show fullsize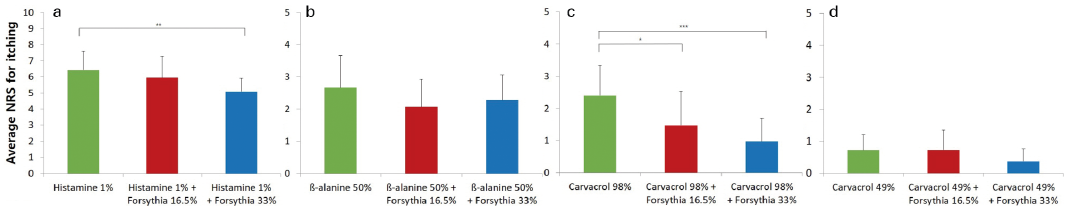 Click to show fullsize
Click to show fullsize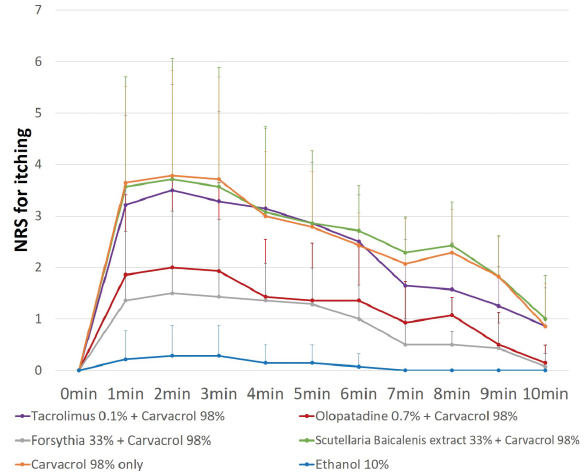 Click to show fullsize
Click to show fullsize Click to show fullsize
Click to show fullsize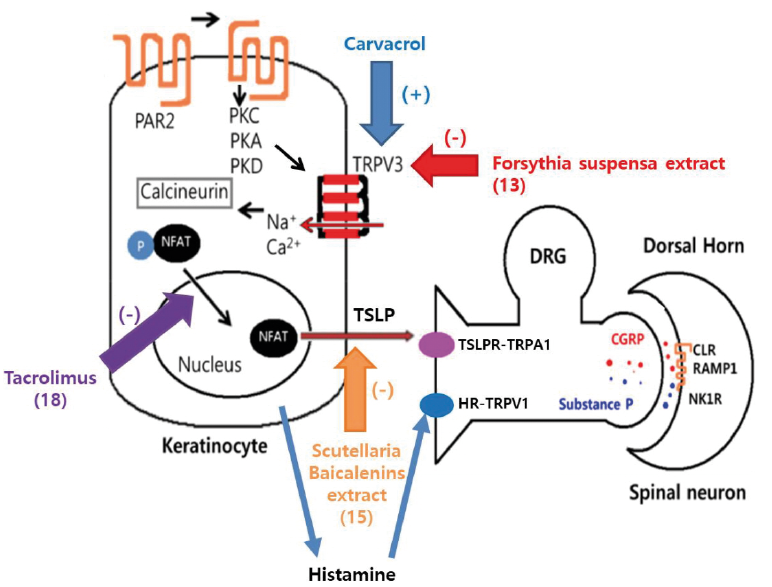 Click to show fullsize
Click to show fullsize