
From the 1Department of Sports Medicine, Medical Clinic, University Hospital Heidelberg and 2Clinic of Orthopedics and Traumatology, University Hospital Heidelberg, Heidelberg, Germany
Objective: Recovery of the quadriceps femoris muscle after anterior ligament reconstruction is impaired. The aim of this study was to investigate satellite cell content and function of the vastus later-alis muscle after anterior ligament reconstruction.
Methods: Biopsies were obtained from the vastus lateralis muscle of 16 recreational athletes immediately before and again 12 weeks after anterior ligament reconstruction. Total satellite cell number (Pax7+), activated (Pax7+/MyoD+), differentiating (Pax7–/MyoD+), and apoptotic (Pax7+/TUNEL+) satel-lite cells, myofibers expressing myosin heavy chain (MHC) I and II, and neonatal MHC (MHCneo) were determined immunohistochemically.
Results: After anterior ligament reconstruction, the number of apoptotic satellite cells was significantly (p = 0.019) increased, concomitant with a significant (p < 0.001) decrease in total satellite cell number, with no change in activated and differentiating satellite cell number. MHCneo+ myofibers tended towards an increase.
Conclusion: Satellite cell apoptosis and the reduction in the satellite cell pool might provide an explanation for prolonged quadriceps muscle atrophy after anterior ligament reconstruction.
Key words: satellite cells; apoptosis; muscle regeneration; developmental myosin heavy chain; muscular atrophy; quadriceps muscle.
Accepted Jan 18, 2021; Epub ahead of print Feb 11, 2021
J Rehabil Med 2021; 53: jrm00153
Correspondence address: Birgit Friedmann-Bette, Department of Sports Medicine, Medical Clinic, University Hospital Heidelberg, Im Neuenheimer Feld 410, 69120 Heidelberg, Germany. E-mail: Birgit.Friedmann-Bette@med.uni-heidelberg.de
Doi: 10.2340/16501977-2794
Protracted muscle atrophy is common after anterior ligament reconstruction, even if athletes adhere to a structured rehabilitation programme. Satellite cells, the stem cells of skeletal muscle, play an important role in recovery of an atrophied muscle. Exercise can activate satellite cells, induce their proliferation, and probably also differentiation of these stem cells. The current study evaluated satellite cell content and function in biopsies from the vastus lateralis muscle of 16 recreational athletes immediately before and 12 weeks after anterior ligament reconstruction. After anterior ligament reconstruction, an increased number of satellite cells showed signs of apoptosis (cell death). Furthermore, total satellite cell number was decreased, with no change in the numbers of activated and differentiating satellite cells. The number of regenerating myofibers expressing neonatal myosin tended to increase. In conclusion, satellite cell apoptosis and the reduced satellite cell number might provide an explanation for the impaired muscle recovery after anterior ligament reconstruction.
Protracted atrophy and weakness of the quadriceps muscle are common after anterior cruciate ligament (ACL) injury and/or anterior cruciate ligament reconstruction (ACL-R), even if the patients undergo guided rehabilitation programmes (1–4). Muscle recovery is compromised due to negative changes in the knee extensor muscles, most likely due to impaired neuromuscular function (5), post-surgery inflammation (6) and immobilization (7). After ACL injury, fibrogenic alterations were observed in biopsies obtained from the vastus lateralis muscle of the injured leg (8, 9). Satellite cell (SC) abundance was also reduced compared with biopsies taken from the vastus lateralis muscle of the uninjured leg (8, 10). Furthermore, there was a surprising lack of increase in SC number after regular rehabilitation training (10), as well as after 12 weeks of supervised quadriceps strength training during rehabilitation after ACL-R (3).
SCs play an important role in skeletal muscle growth and regeneration (11, 12). Increases in SC number occur after 11–12 weeks of quadriceps strength training in healthy subjects (13–15). The role of SCs in atrophy of human skeletal muscle, however, has scarcely been investigated. In the very few studies on the role of SCs in atrophy of human skeletal muscle (8, 10, 16), loss of SCs with atrophy is not a consistent finding. However, there is some evidence that a particularly severe atrophic environment, as is found, for example, after severe burn injury, has a negative impact on SC number and SC function and can induce SC apoptosis (17). With regard to findings in animal studies, it has been hypothesized that muscle wasting in old age (sarcopaenia) could at least partly be explained by SC dysfunction with increased SC apoptosis due to chronic low-grade systemic inflammation (18). The significantly reduced SC number in biopsies from the vastus lateralis muscle after ACL injury (8, 10) and the lack of increase in SC number after resumption of muscular training after ACL-R (3, 10) suggest that ACL injury and/or ACL-R with quadriceps tendon or semitendinosus tendon autografts, respectively, might generate a severe atrophic environment with negative effects on SC number and function.
The primary aim of this study was to further investigate the effects of ACL-R on SCs and to determine whether the previously described reduction in SC number might be due to SC apoptosis. The study analys-ed muscle biopsies from the vastus lateralis muscle of recreational athletes immediately before ACL surgery and again after 12 weeks of early rehabilitation.
Participants
A total of 16 recreational athletes who underwent standardized ACL-R (mean age 26 years (standard deviation (SD) 4), mean height 180.0 cm (SD 7.0), mean weight 83.3 kg (SD 13 kg), 15 men, 1 woman, volunteered to participate in the study. Immediately before starting the ACL surgery and again after finishing the regularly scheduled 12 weeks’ early rehabilitation, biopsies were obtained from the vastus lateralis muscle of the injured leg. Eleven of the athletes also participated in our study on strength training effects on muscular regeneration after ACL-R (3). The study was approved by the ethics committee of the local Medical Faculty and was conducted in accordance with the principles of the Declaration of Helsinki. Each subject provided written informed consent to participate.
Early rehabilitation
During the 12-week period of early rehabilitation, the injured leg was not completely immobilized, but subjected to successively increasing exercises and load bearing with restricted range of motion (ROM) under the control of physiotherapists, the progress being adapted with regard to the swelling of the knee and pain-free ROM. The rehabilitation programme aimed to reach full ROM and load bearing after 12 weeks, to enable the patients to participate in strength training and coordination exercises.
Muscle biopsy sampling
Immediately before starting the ACL surgery and again after the 12-week early rehabilitation period, muscle biopsy samples were taken from the same region at mid-thigh level of the vastus lateralis muscle of the injured leg under local anaesthesia, using the Bergström technique (19). In detail, the skin, subcutaneous tissue and outer fascia were carefully anaesthetized with 3 ml prilocaine (1%) using a 23G needle. An approximately 2-cm skin incision was performed as well as a small incision of the fascia (approximately 5 mm) so that the biopsy needle could be inserted into the vastus lateralis muscle at a depth of approximately 2–3 cm. Under suction, a mean of 50–100 mg of muscle tissue was obtained. The muscle tissue was immediately freed from blood and visible connective tissue, rapidly frozen in isopentane cooled by liquid nitrogen and subsequently stored at –80°C. To avoid residual effects from preceding biopsies, biopsy sites were spaced approximately 1 cm apart in the longitudinal direction from distal to proximal.
Muscle immunohistochemistry
All biopsies were assigned a random unique identification number, thereby blinding the investigator to subject identity and time point. However, simultaneous analysis of the biopsies from each subject before and after early rehabilitation in the same staining procedure was ensured. Immunofluorescent analyses were performed on serial sections for SC number, activated SCs, apoptotic SCs, fibre type distribution, and myofibers expressing neonatal myosin (MHCneo). For all analyses, transverse sections (10 µm) were cut in a cryostat (–20°C) mounted on Super Frost Plus slides (ThermoFisher Scientific, Waltham, MA, USA) and allowed to dry for 20 min. Sections were then fixed in 4% paraformaldehyde (PFA, Roti-Histofix 4%, catalogue number P087; Carl Roth, Karlsruhe, Germany) for 10 min. For all further procedures, except the analysis for apoptotic SCs, sections were then washed 3 × 5 min in phosphate-buffered saline (PBS) and blocked for 10 min at room temperature (RT) in PBS containing 0.5% Triton (Triton X-100, catalogue number 108643; Merck Millipore, Billerica, MA) and 1% bovine serum albumin (BSA, Albumin Bovine Fraction V, catalogue number 11922; Serva Electrophoresis, Heidelberg, Germany).
Satellite cell number
For determination of the SC number, staining of Pax7, laminin, type II myosin and nuclei was performed on the same section. Sections were incubated overnight at RT with the primary antibodies anti-Pax7 (1:500; catalogue number MAB1675, Bio-Techne (R&D Systems), Minneapolis, MN, USA), anti-laminin (1:1,000, catalogue number ab11575, Abcam, Cambridge, UK) and anti-Fast Myosin Skeletal Heavy Chain (1:1,000, catalogue number ab91506, Abcam). The next day, the sections were washed in PBS, followed by incubation in appropriate secondary antibodies: Alexa-fluor 488 goat-antimouse (Pax7; 1:1,000, catalogue number A.11029, ThermoFisher (Invitrogen)) and Alexa-fluor 568 goat-antirabbit (Laminin and MHC II; 1:1,000, catalogue number A.11036, ThermoFisher (Invitrogen)) for 2 h at RT. Then, the sections were washed in PBS and nuclei were visualized with bisBenzimide (Hoechst 33258, catalogue number B2883, Sigma-Aldrich, St Louis, MO, USA). Finally, sections were washed in PBS, allowed to dry completely in the dark and then fixed with FluorSave Reagent (catalogue number 345789, Merck Millipore) at 4°C for 24 h. Staining specificity was confirmed using appropriate negative controls.
Activated satellite cells
For analysis of myogenic activation, sections were incubated overnight at RT with primary antibodies against Pax7 (1:500; catalogue number MAB1675, Bio-Techne (R&D Systems)) and against MyoD (1:500; catalogue number ab133627, Abcam). The secondary antibodies Alexa-fluor 488 goat-antimouse and Alexa-fluor 568 goat-antirabbit were used for visualisation of Pax7 and for MyoD, respectively, and the staining procedure was finished, as described previously.
Apoptotic satellite cells
For determination of SC apoptosis, terminal deoxynucleotidyl transferase-mediated dUTP nick end-labelling (TUNEL) and staining of Pax7 were conducted on the same section. After the slides were fixed in 4% PFA as described above, the sections were blocked, permeabilized, washed and incubated in the In Situ Cell Death Detection Kit (catalogue number 11684817910 Roche, Sigma-Aldrich) according to the manufacturer’s instructions. Afterwards, the slides were washed 3 × 5 min in PBS and blocked for 10 min at RT in PBS containing 0.5% Triton and 1% BSA. The sections were then incubated overnight at RT with the primary antibody against Pax7 (1:500; catalogue number MAB1675, Bio-Techne (R&D Systems)). The second-ary antibody Alexa-fluor 568 goat-antimouse (1:1,000, cata-logue number A.11031, ThermoFisher (Invitrogen)) was used for visualization of Pax7 and the staining procedure was finished, as described previously. Staining specificity was confirmed using appropriate negative and positive controls.
Fibre-type distribution
To enable categorization into type I, type II or hybrid fibres, sections were incubated overnight at RT with the primary antibodies anti-Myosin (Skeletal, Slow; MHC I, 1:6,000, catalogue number M8421, Sigma Aldrich) and anti-Fast Myosin Skeletal Heavy chain (MHC II, 1:1,000, catalogue number ab91506, Abcam). Secondary antibodies Alexa-fluor 488 goat-antimouse and Alexa-fluor 568 goat-antirabbit were used for visualization of MHC I and MHCII, respectively, and the staining procedure was finished as described above.
Myofibers expressing MHCneo
Regeneration/remodelling was assessed by staining of sections for neonatal MHC. Sections were incubated overnight with the primary antibodies anti-MHCneo (1:75, NCL-MHCn, Leica Biosystems, Wetzlar, Germany), anti-Laminin (1:1,000, catalogue number ab11575, Abcam) and anti-Fast Myosin Skeletal Heavy Chain (catalogue number ab91506, Abcam). The second-ary antibody Alexa-fluor 488 goat-antimouse was used for visualization of MHCneo and Alexa-fluor 568 goat-antirabbit for laminin as well as for MHC II. The staining procedure was finished as described above.
Image analyses
Images were obtained at × 20 and at × 40 magnification using the fluorescence microscope Axio Observer and the appropriate software programme Zen (Carl Zeiss, Oberkochen, Germany). The analysed mean tissue surface area was 1.78 mm2 (SD 1.29) before and 2.12 mm2 (SD 0.95) after ACL-R and early rehabilitation; 265 myofibers (SD 225) and 259 myofibers (SD 174) per section were analysed, respectively. The criterion for a SC was simultaneous Pax7 and myonucleus (bisBenzimide Hoechst 33258) staining within the myofiber (sublaminar). The number of Pax7 positive (Pax7+) cells (SCs) associated with MHC I+ (type I) or MHC II+ (type II) fibres was quantified and the number of SCs divided by the total number of myofibers was categorized as SCs per fibre. Activat-ed SCs were characterized by simultaneous MyoD, Pax7 and bisBenzimide staining (MyoD+/Pax7+/bisBenzimide). MyoD and bisBenzimide staining in Pax7 negative cells (MyoD+/Pax7–/bisBenzimide) was regarded as indicating differentiating SCs. The numbers of activated SCs and differentiating SCs were each divided by the total number of myofibers. Apoptotic SCs were classified as Pax7+/TUNEL+/bisBenzimide. The number of apoptotic SCs was expressed relative to total SC number. The number of MHCneo positive fibres was related to total fibre number from the section. FCSAs were measured in images obtained at ×20 magnification using the software programme Axio Vision (Carl Zeiss, Oberkochen, Germany). Only fibres cut perpendicular to their longitudinal axis were used for determination of fibre size.
Statistical analyses
All statistical tests were processed using SPSS 24.0 software for Windows (SPSS Inc., Chicago, IL, USA). Paired samples Student’s t-tests were performed to compare the results obtain-ed before vs 12 weeks after ACL-R. The normal distribution of all data was checked using the Shapiro-Wilks test. This test failed for the number of fibres expressing MHCneo and for the number of activated SCs. For these data, the Wilcoxon signed-rank test was applied. To investigate the relationship between selected parameters, Pearson product moment correlations were calculated. For all analyses, a value of p < 0.05, set a priori, was considered to represent statistical significance. The figures were created using GraphPad Prism 7.0 (GraphPad Software Inc., La Jolla, CA, USA).
Data are presented as mean (SD).
Satellite cells
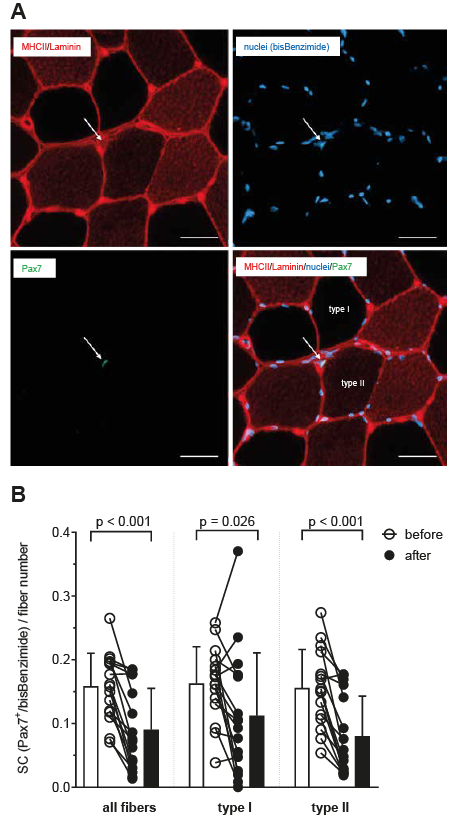
Significant decreases were observed for the SC number (Pax7+/bisBenzimide) in relation to total myofiber number (p < 0.001) as well as related to type I (p = 0.026) and to type II fibres (p < 0.001) 12 weeks after ACL-R and early rehabilitation (Fig. 1). SC number per cross-sectional area decreased significantly (p = 0.0013) from 26 SCs per mm2 (SD16) to 15 SCs per mm2 (SD 11).
Fig. 1. Identification of satellite cells. (A) Representative muscle cross-section stained for myosin heavy chain (MHC) II (red) and laminin (red), for nuclei (bisBenzimide, blue), and for Pax7 (green). One satellite cell (SC) is identified, related to type II fibre. Scale bar: 50 µm. (B) SC content presented as SC number related to total fibre number as well as to type I and type II fibres before and after anterior cruciate ligament reconstruction (ACL-R) and 12 weeks of early rehabilitation. Individual data-points as well as mean values (standard deviation; SD) are shown.
No significant changes occurred for the number of activated SCs (Pax7+/MyoD+/bisBenzimide) and differentiating SCs (Pax7–/MyoD+/bisBenzimide) in relation to total myofiber number (Fig. 2).
Fig. 2. Identification of activated and differentiating satellite cells (SC). Representative muscle cross-section stained for MyoD (red), nuclei (bisBenzimide, blue) and Pax7 (green). (A) Two SC are identified, one of them activated (MyoD+/Pax7+/bisBenzimide). (B) Two SCs are identified, one of them differentiating (MyoD+/Pax7-/bisBenzimide). Scale bar: 20 µm. (C) Individual data-points and mean values (standard deviation; SD) of activated (Pax7+/Myo+) and differentiating (Pax7-/MyoD+) SCs per myofiber before and after anterior cruciate ligament reconstruction (ACL-R) and 12 weeks of early rehabilitation. (D) Distribution of SC number (Pax7+/MyoD-), activated SCs (Pax7+/Myo+), and differentiating SCs (Pax7–/MyoD+) before and after ACL-R and 12 weeks of early rehabilitation.
The number of apoptotic SCs (Pax7+/TUNEL+/bisBenzimide) related to total SC number (Pax7+/bisBenzimide) was significantly (p = 0.019) increased 12 weeks after ACL-R and early rehabilitation. Twelve weeks after ACL-R, approximately 35% of all SCs were TUNEL-positive compared with approximately 1% before ACL surgery (Fig. 3).
The changes in numbers of SC and apoptotic SCs were not significantly correlated with age, sex, or time between injury and surgery.
Fig. 3. Identification of apoptotic satellite cells (SC). Representative muscle cross-sections showing myonuclear apoptosis (A) before anterior cruciate ligament reconstruction (ACL-R) and (B) 12 weeks after ACL-R and early rehabilitation in the same individual. (A) Three satellite cells (SCs) are shown (arrows), one of them apoptotic. (B) Two SCs are shown (arrows), both of them apoptotic. TUNEL (green), nuclei (bisBenzimide, blue), Pax7 (red), TUNEL/Pax 7/bisBenzimide (white). Scale bar: 50 µm. (C) Incidence of SC apoptosis presented as number of apoptotic SCs (Pax7+/TUNEL+/bisBenzimide) related to total number of SCs (Pax7+/bisBenzimide) before and after ACL-R and 12 weeks of early rehabilitation. Individual data points as well as mean values (standard deviation; SD) are shown.
Fibre type distribution and fibre cross-sectional areas
No significant changes in the proportion of type I fibres (39.1% (SD 15.8) vs 40.7% (SD 19.5), type II fibres (41.6% (SD 13.7) vs 43.0% (SD14.5), and hybrid fibres (19.4% (SD 6.3) vs16.3% (SD 11.0)) were found 12 weeks after ACL-R and early rehabilitation. FCSA of type I and of type II fibres tended towards a decrease 12 weeks after ACL-R and early rehabilitation by a mean of 16% and 12%; however, the level of significance was not reached (Fig. 4).
Fig. 4. Fibre type distribution and fibre cross-sectional areas. (A) Myofibers expressing myosin heavy chain (MHC) I (red, type I), MHC II (yellow, type II), and MHC I/MHC II (orange, hybrid fibres). Scale bar: 50 µm. (B) Cross-sectional areas of the different fibre types before and after anterior cruciate ligament reconstruction (ACL-R) and 12 weeks of early rehabilitation. Individual data points as well as mean values (standard deviation; SD) are shown.
Regenerative myofibers (expressing MHCneo)
Related to total myofiber number, the number of MHCneo positive fibres tended to increase by a mean of 80%; however, due to considerable inter-individual variability, the level of significance was not reached (Fig. 5). All but one myofiber expressing MHCneo were also MHC II positive (MHCneo+/MHC II+).
Fig. 5. Fibres expressing neonatal myosin heavy chain (MHC). (A) Representative muscle cross-section stained for MHC II (red) and laminin (red), for nuclei (bisBenzimide, blue) and for neonatal MHC (MHCneo, green). One fibre expressing neonatal MHC is shown, also staining for MHC II. Scale bar: 50 µm. (B) Individual data-points and mean values (standard deviation; SD) of MHCneo+ fibres related to total fibre number before and after anterior cruciate ligament reconstruction (ACL-R) and 12 weeks of early rehabilitation.
Disuse atrophy is a common finding after ACL-R. This study found a significant decrease in the number of SCs in biopsies obtained from the vastus lateralis muscle 12 weeks after ACL-R and early rehabilitation compared with the analyses performed in biopsies obtained immediately before ACL surgery, together with a trend towards reduction in the FCSAs of all fibre types. The main new finding, however, was a significant increase in SC apoptosis as a potential cause for the compromised SC pool after ACL-R.
SCs are regarded as the main candidates that play an essential role in muscle repair and remodelling (11, 20). Some studies investigating the loss of muscle mass have reported a reduced number of SCs during immobilization in a bedrest study (21), during ageing (22–24), and in spinal cord injury (22). The significant reduction in number of SCs observed in the vastus lateralis muscle 12 weeks after ACL-R in the current study, as well as in the injured compared with the uninjured leg before ACL-R in other studies (8, 10) might, therefore, provide an explanation for the protracted quadriceps muscle atrophy after ACL-tear and ACL-R, respectively. There is some evidence that the accretion of new myonuclei from activated SCs after resistance exercise is of importance for the development of muscle hypertrophy (11). A decreased pool of SCs probably has a negative impact on the regain of muscle mass during rehabilitation after ACL-R because less new myonuclei can be provided from activated SCs in response to exercise. However, not only the decreased pool of SCs, but also reduced SC function might impair muscle recovery after ACL-R. In contrast to the common finding of an increase in number of SCs after several weeks of strength training (13–15), there was a lack of increase in the number of SCs after resumption of strength training (3) or after a rehabilitation programme that enabled the athletes to resume sport-specific drills (10). Further evidence of impaired SC function after ACL-R is provided by the observation of increased apoptosis of SCs in the current study, 12 weeks after ACL-R and early rehabilitation, when the athletes had reached full ROM and weight-bearing of the injured knee and were cleared by the orthopaedic surgeon for resumption of strength training. An increased frequency of myonuclear apoptosis (TUNEL+ myonuclei) was also described in biopsies obtained from the vastus lateralis muscle of athletes who had had an ACL tear within 2 months before the biopsies were taken; however, the authors did not show SC apoptosis (8). The degree of apoptosis across all cell types appeared to be lower in that investigation than in the current study, where the subjects first had an ACL injury and then were subjected to ACL-R, post-surgery immobilization and 12 weeks of early rehabilitation. It can be speculated that all these procedures caused a higher myonuclear turnover. However, further investigation is needed regarding this unexpected finding.
There are few studies on apoptosis of SCs in human skeletal muscle. No apoptosis of SCs was found after 14 days of immobilization due to unilateral whole-leg casting in healthy young and old individuals (25). In contrast, an increased number of apoptotic SCs was observed in the soleus muscle of mice after a 14-day hindlimb unloading procedure. Electrical stimulation of the suspended muscles led to a significant reduction in apoptosis of SC; how-ever, apoptosis of SC was still increased compared with the findings in a control group of weight-bearing mice (26). With regard to these findings in an animal study, immobilization might cause SC apoptosis, which can probably be counteracted by muscle contractions induc-ed by electrical stimulation. In human skeletal muscle, increased SC apoptosis was reported in children with burn injury 17 days (SD 5) post-burn, when the activity level was characterized by physical and occupational therapy sessions (17). These results are in accordance with the findings of an animal study after scald-burn injury (27). It is very likely that alterations in the SC niche, the microenvironment surrounding the SCs, are the cause of SC apoptosis and/or SC dysregulation observed after severe burn injury. There is growing evidence that, besides the well-known effects of stimulatory and inhibitory growth factors, extracellular matrix components are essential mediators in the SC niche (12, 28). In particular, interactions between SCs and fibroblasts, which are considered the main contributor to the extracellular matrix of skeletal muscle, seem to be of great importance for muscle regeneration (29). With regard to expansion of the extracellular matrix with increased collagen deposition observed concurrently with a reduced number of SCs in the vastus lateralis muscle after ACL injury (8), the cross-talk between fibroblasts and SCs might be disturbed after ACL injury. The expansion of the extracellular matrix also described in the vastus lateralis muscle of severely burned children, together with the reduced SC pool and increased SC apoptosis (17) points in the same direction.
In the current study, there was a tendency towards an increase in myofibers expressing MHCneo 12 weeks after ACL-R and early rehabilitation, although with a wide inter-individual variation. This finding is in accordance with the observation of a significantly increased number of myofibers containing embryonic myosin heavy chain, another developmental myosin heavy chain, after severe burn injury (17, 27). The developmental myosin heavy chains are transiently expressed during embryonic and foetal development; their re-expression in the adult skeletal muscle is regarded as indicative of degeneration/regeneration events in the pathological human skeletal muscle (e.g. in myopathies or polymyositis) and in regenerative processes after severe muscle damage in animal studies (30). In the regenerating myofiber, MHC expression switches from developmental MHC to fast MHCs independently from innervation (30) providing an explanation for the concomitant expression of MHCneo and MHC II in all but one of the regenerative myofibers in the current study. The switch to slow myosin is controlled by slow nerve activity (30), which is low during immobilization. The numbers of activated and differentiating SCs remained unchanged 12 weeks after ACL-R and early rehabilitation in the current study. In contrast, a greatly increased number of activated SCs was observed in the vastus lateralis muscle of severely burned children, but this increase was interpreted as a sign for inefficient myogenesis due to dysregulated terminal activation of SCs in the presence of systemic factors induced by burn injury (17).
Study limitations
The current study has several limitations. The causes of the observed increase in SC apoptosis 12 weeks after ACL-R and early rehabilitation can only be speculated; for example, it was not possible to prove increased fibrotic alterations in the muscle biopsies obtained at that time-point, because reliable staining to assess fibrosis could not be performed. Another limitation is that no biopsies were obtained from the contralateral leg. It is not known if the reduction in number of SCs and the increased SC apoptosis resulted from ACL-R and the special physical treatment during early rehabilitation or from a general reduction in physical activity. There are only a few studies on SC apoptosis in human skeletal muscle and, to our best knowledge, no publications on SC apoptosis in muscle tissue of subjects undergoing ACL-R. Therefore, these novel results could only be compared with observations in animal studies or with findings in the muscles of severely burned children. Of course, such comparison can only be made with great caution. In particular, for comparison with the ob-servations of the muscles of severely burned children, the differences in the muscular stressors and the patients investigated must be considered.
Conclusion
In conclusion, the significantly increased SC apoptosis observed in the current study concomitantly with a significantly reduced SC pool and regenerative myofibers 12 weeks after ACL-R and early rehabilitation adds further evidence to the observation of an impaired muscle quality several weeks after ACL-R. Although the athletes were cleared by the orthopaedic surgeon to resume strength training and sport-specific drills, their quadriceps femoris muscle was probably not ready to respond to the training stimuli with development of an adequate muscle hypertrophy and increase in muscle strength.
The study was funded by the Dietmar Hopp Foundation (Project 23011193). The authors thank Dr Nikolaus Streich, Dr Jan-Paul Flacke, Steffi Kraushaar, and Axel Schöffel for their assistance in the study.
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize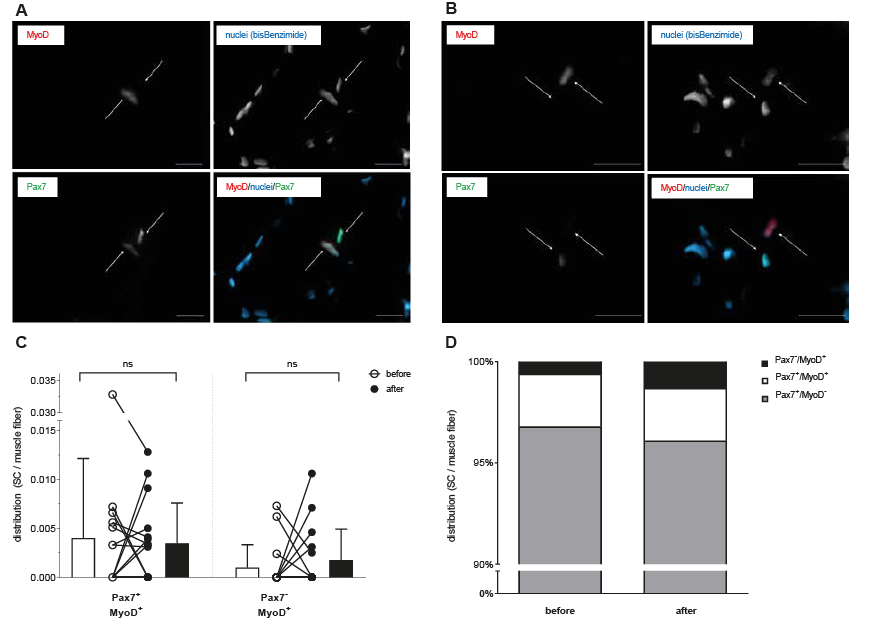 Click to show fullsize
Click to show fullsize Click to show fullsize
Click to show fullsize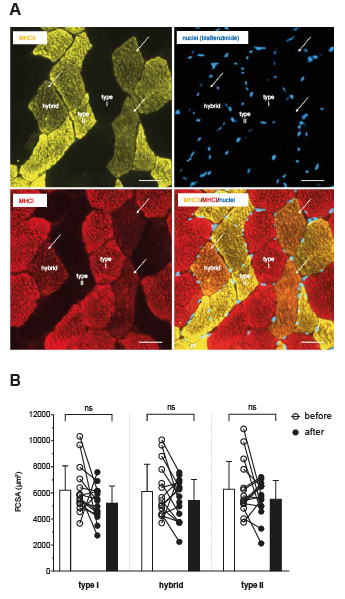 Click to show fullsize
Click to show fullsize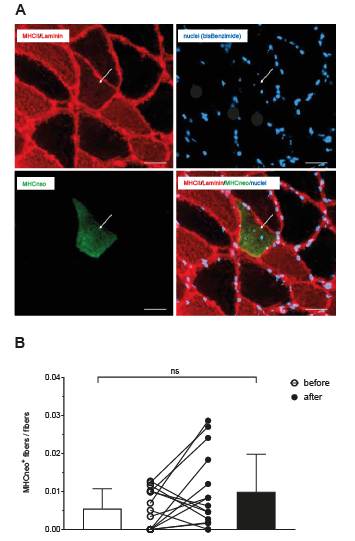 Click to show fullsize
Click to show fullsize