


Céline Girard1, Laure Ottomani2, Jacques Ducos2, Olivier Dereure1, Marie J. Carles3 and Bernard Guillot1
1Service de Dermatologie, Hopital Saint Eloi, CHU Montpellier, Laboratoire de Virologie, 2Hopital Lapeyronie, CHU Montpellier, Montpellier, and 3CHU de Nimes, Nimes, France
Human herpes virus 8 infection is the primary and necessary factor in the development of Kaposi’s sarcoma, but is not sufficient per se to trigger the onset of the disease. In order to search for virological cofactors associated with the occurrence of the disease, we investigated the prevalence of active infection by two newly discovered viruses, hepatitis G virus and TT virus, among patients with classical Kaposi’s sarcoma. Serum of 24 patients with Mediterranean Kaposi’s sarcoma was investigated using polymerase chain reaction and compared with that of 68 healthy subjects. Cutaneous samples from patients with Kaposi’s sarcoma and healthy subjects were investigated for TT virus DNA. No patient had serum markers for hepatitis G virus. TT virus DNA was present in the serum of 21/24 (87.5%) patients and 32/68 (47%) controls (p = 0.002). TT virus DNA was present in the lesional skin of 5/18 patients with Kaposi’s sarcoma (27.7%), but not in the skin of controls. TT virus might play a role as a cofactor in the clinical emergence of Kaposi’s sarcoma in patients infected with Human herpes virus 8, perhaps by immunosuppressive effects or by a common transmission pathway for these two viruses. Key words: Kaposi’s sarcoma; HHV-8; hepatitis G virus; TT virus.
(Accepted August 29, 2006.)
Acta Derm Venereol 2007; 87: 14–17.
Céline Girard, Service de Dermatologie, Hopital Saint Eloi, CHU Montpellier, FR-34280, Montpellier, France. E-mail: girard-drouard@tele2.fr
In 1994, Chang et al. (1) identified a new human herpes virus called HHV-8, which was associated with all epidemiological subsets of Kaposi’s sarcoma (KS) and always present in patients’ involved skin. HHV-8 can be transmitted sexually or non-sexually. A maternal–infant transmission has also been described (2). In a recent study, Brambilla et al. (3) studied the prevalence of HHV-8 seropositivity among the sexual partners of patients with classical KS: 44% were seropositive for HHV-8, but none had developed the disease, suggesting that HHV-8 infection alone is not sufficient per se to trigger the onset of the disease. Other cofactors, such as virological, environmental, genetic, metabolic or iatrogenic factors, might thus be involved in the development of KS, particularly through immune system dysfunction. In order to search for virological cofactors associated with the occurrence of the disease, we investigated the prevalence of active infection by two newly discovered viruses, hepatitis G virus C (GBV-C) and TTV (Torque teno virus), among patients with classical KS. The flavivirus GBV-C, (also designated hepatitis G virus) was identified in a search for hepatitis viruses, but no disease is currently known to be associated with it. The TT virus was first isolated in 1997 in a Japanese patient with acute post-transfusion hepatitis and it was thus considered to be transfusion-transmitted and hepatotropic. The virus was named TT, both in reference to the initials of the first patient with detectable TTV DNA in serum and as an abbreviation of “transfusion-transmitted”. More recently however, taxonomists have further proposed that the full name for TTV be Torque teno virus, within the genus Anellovirus (4). Several attempts have been made to link TTV to the aetiology of a specific disease, but no significant association with human pathology has yet been reported.
GBV-C replicates in lymphocytes; inhibits the replication of human immunodeficiency virus (HIV) in vitro; and has been associated with improved survival, improved AIDS-free survival, higher CD4 (+) T-cell counts, and lower HIV viral loads in HIV-infected people compared with people infected with HIV but not GBV-C (5).
The frequent identification of TTV in peripheral mononuclear blood cells suggests that the virus replicates in lymphoid cells and contributes to lymphocyte imbalances with consequent immunosuppressive effects (6).
Based on these recent findings, GBV-C and TTV may be ideal candidates for use as immunomodulator agents. The aim of this study was to investigate the presence of these viruses in KS.
MaterialS and methods
Twenty-four patients (21 men and 3 women; age range 29–97 years; mean 62.9 years), with Mediterranean KS were enrolled in the study. HIV infection was ruled out by serological testing (enzyme-linked immunoassay (ELISA) test: Genscreen Plus HIV Ag-Ab, BioRad, France). In each case, KS was confirmed by histological examination and serological testing for HHV-8 was performed (immunofluorescent assay, Biotrin, Dublin, Ireland) based on the latency-associated nuclear antigen of the virus.
GBV-C RNA was searched for in serum using a reverse transcription-polymerase chain reaction (RT-PCR) method. Briefly, RNA was extracted from serum using QIAamp Viral RNA Minikit (QIAGEN GmbH, Hilden, Germany) and RT-PCR was performed with AC1S 7AS primers (AC1S: 5'-AGG GTT SGW NGG TSG TAA ATC C-3'; 7AS: 5'-CAA GAG MGR CAT TGA AGR GCG A-3').
TTV DNA was searched for using a PCR method. Total genomic DNA was extracted from serum using the QIAamp DNA Minikit (QIAGEN GmbH, Hilden, Germany) and amplified by PCR with a set of specific primers (T801 T935) (7). Amplicons were bi-directionally sequenced using the "OpenGene" sequencing system (7), and the resulting sequences were queried against a library of known genotypes.
As a control group, 68 blood donors matched for age and gender were tested for both serum GBV-C RNA and TTV DNA.
Samples of lesional and normal skin from 18 patients with KS were investigated (the other patients refused the skin biopsy), HHV-8 DNA was searched for in both lesional and non-lesional skin using a specific nested technique, as previously described (1), as to the presence of TTV DNA using the same amplification method than in serum.
TTV DNA was also searched in cutaneous samples obtained from 7 healthy subjects following aesthetic surgery.
The prevalence of antibodies to HHV-8 and positivity for HHV-8 DNA, GBV-C RNA and TTV DNA was defined as the number of positive individuals per total number of those tested. A χ2 test was used to analyse categorical variables. P-values lower than 0.05 were considered to indicate statistical significance.
Results
The serological test for HHV-8 was positive in all patients. No patient had serum markers for GBV-C. TTV DNA was present in the serum in 21/24 patients (87.5%) (Fig. 1). One female and two male patients were negative for TTV. Sequencing of the amplicons confirmed TTV DNA.
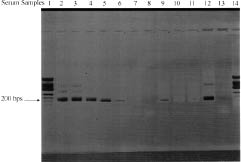
Fig. 1. Sera samples of patients with classical Kaposi’s sarcoma. DNA was extracted and reverse transcription-polymerase chain reaction was performed as described in the Materials and Methods section. The gel shows a 200 bps band for TTV DNA (lines 2, 3, 4, 5, 6 and 9). Lines 7, 8, 10 and 11 correspond to TTV negative patients. Lines 1 and 14 are molecular weight markers (no. 9, Eurogentec, Belgium), line 12 is a positive control and line 13 a negative control (sterile water).
TTV DNA was present in 32/68 (47%) control subjects and GBV-C RNA was present in the serum of 3/68 (4.4%).
HHV-8 DNA was detected in lesional skin in all patients (18/18) and in non-lesional skin in 3/18 patients (16.6%). In patients with KS, TTV DNA was present in lesional skin in 5/18 patients (27.7%) and in normal skin in 3/18 patients (16.6%). The three positive patients with TTV DNA in normal skin had TTV DNA in lesional skin as well. None of seven cutaneous samples from control subjects disclosed TTV DNA.
Discussion
The association between KS and a number of viruses including cytomegalovirus, human herpes virus type 6 and 7, human papilloma virus and Epstein Barr virus has been researched without any convincing results (8). On the other hand, over 95% of KS lesions, regardless of their source or clinical subtype, have been found to be infected with HHV-8 (2). HHV-8 can be transmitted sexually (9) and by other means. Maternal–infant transmission, whether during labour and delivery or transplacentally, accounts for a portion of HHV-8 infections (10). The other exact routes of transmission are not known, although HHV-8 has been detected in both saliva (11) and semen (12) from infected individuals. Infection by hepatitis G virus and TTV concomitant with KS has never been studied to our knowledge.
Hepatitis G virus or GBV-C virus is likely to be transmitted by blood exposure more frequently than by sexual route. The prevalence of GBV-C RNA in the serum of a population of healthy French blood donors is rated at 4.2%, according to Loiseau et al. (13). None of our patients had serum markers of active infection by GBV-C, suggesting that this virus is not involved in the occurrence of KS.
The prevalence of TTV in human populations has been continuously re-evaluated, in direct correlation with the design and optimization of DNA amplification systems (7, 14, 15). The prevalence of TTV DNA presence in serum of the general population from southern France is high, estimated as between 49.5% and 66% (14, 16), which is consistent with the data from our control group (47%). The difference in TTV DNA prevalence in serum between control group and patients with KS was statistically significant (p = 0.002, χ2- test). Moreover, TTV DNA was detected in both non-lesional and lesional skin of patients with KS in 16.6% and 27.7%, respectively, of the investigated samples, whereas it was absent in all samples from the control group.
TTV is a newly described DNA virus, whose involvement in human pathological conditions remains elusive. This virus seems to be transmitted by several modes, including blood exposure, based on the high prevalence of TTV DNA in haemophiliacs and dialysed or transfused patients, and, to a lesser extent, sexual intercourse (17). Conversely, orofaecal transmission might be an important route, as suggested by the high rate of viral excretion in bile (18). In a recent report (19), high loads of TTV were found in the nasal secretions of infants, suggesting that the virus may be airborne. In addition, the recent finding of high levels of TTV in saliva suggests that this biological fluid may be an efficient vehicle for virus transmission (20). In patients with KS infected with HHV-8, TTV might play a role as a cofactor through a common pathway of transmission for these two viruses.
Moreover, TTV may have effects on the host’s immune system and aggravate disease induced by other agents. From this perspective, several studies analysed the role of TTV in human tumours (21–23). An increased detection rate of TTV was demonstrated in gastrointestinal tumours and in breast and lung cancer, as well as in leukaemias and multiple myelomas (24). Indeed, it was demonstrated recently that high TTV loads were associated with severe forms of disease in children hospitalized for acute respiratory diseases (19). In such children, TTV loads showed an inverse correlation with the percentage of circulating total T cells and helper T cells and a direct correlation with the percentage of B cells (6). Peripheral blood cells have been shown to harbour TTV DNA, suggesting that the virus might replicate in lymphoid cells and contribute to lymphocyte imbalances with consequent immunosuppressive effects (6). In this way TTV infection could lead to the clinical emergence of KS in patients infected with HHV-8 by promoting HHV-8 replication and increasing the HHV-8 viral load.
Further studies are necessary to assess the prevalence of TTV DNA in other groups, such as patients with positive HHV-8 serology but no KS lesions, and in other clinical subsets of KS, such as AIDS-associated KS.
The discovery of HHV-8 was a major breakthrough in understanding KS mechanisms but other cofactors are also likely to be involved, as suggested by the low incidence of KS in the HHV-8-infected population. Our findings did not demonstrate an aetiological role for TTV in KS but they certainly raise questions about the significance of TTV in disease pathogenesis and about whether this is mediated via immune system dysfunction. It is justified to pursue the study of TTV infection in humans and to investigate any potential relationship with disease.
Simple prevalence studies in various human populations no longer seem sufficient for this purpose, since it has recently been established that the prevalence of TTV infection is high in healthy populations whenever suitable detection tools are used (14, 16). The measurement of viral loads in KS patients could be one alternative, but the enormous genetic diversity of TTV makes the simultaneous study of different groups technically difficult for the moment.
ACKNOWLEDGEMENTS
This work was supported by a grant from the “Ligue Nationale Contre le Cancer, Comité du Gard”.
REFERENCES
1. Chang Y, Cesarman E, Pessin MS, Lee F, Culpepper J, Knowles DM, et al. Identification of herpesvirus-like DNA sequences in AIDS-associated Kaposi’s sarcoma. Science 1994; 266: 1865–1869.
2. Antman K, Chang Y. Kaposi’s sarcoma. N Engl J Med 2000; 342: 1027–1038.
3. Brambilla L, Boneschi V, Ferrucci S, Taglioni M, Berti E. Human herpesvirus-8 infection among heterosexual partners of patients with classical Kaposi’s sarcoma. Br J Dermatol 2000; 143: 1021–1025.
4. Hino S. TTV, a new human virus with single stranded circular DNA genome. Rev Med Virol 2002; 12: 151–158.
5. Williams CF, Klinzman D, Yamashita TE, Xiang J, Polgreen PM, Rinaldo C, et al. Persistent GB virus C infection and survival in HIV-infected men. N Engl J Med 2004; 350: 981–990.
6. Maggi F, Pifferi M, Tempestini E, Fornai C, Lanini L, Andreoli E, et al. TT virus loads and lymphocyte subpopulations in children with acute respiratory diseases. J Virol 2003; 77: 9081–9083.
7. Takahashi K, Hoshino H, Ohta Y, Yoshida N, Mishiro S. Very high prevalence of TT virus (TTV) infection in general population of Japan revealed by a new set of PCR primers. Hepatol Res 1998; 12: 233–239.
8. Drago F, Raineri E, Rebora A. Non-AIDS-related Kaposi sarcoma tissues do not contain DNA sequences of HHV-6, HHV-7, Epstein-Barr virus, cytomegalovirus and HSV. Acta Derm Venereol 1998; 78: 485.
9. Martin JN, Ganem DE, Osmond DH, Page-Shafer KA, Macrae D, Kedes DH. Sexual transmission and the natural history of human herpesvirus 8 infection. N Engl J Med 1998; 338: 948–954.
10. Mayama S, Cuevas LE, Sheldon J, Omar OH, Smith DH, Okong P, et al.Prevalence and transmission of Kaposi’s sarcoma-associated herpesvirus (human herpesvirus 8) in Ugandan children and adolescents. Int J Cancer 1998; 77: 817–820.
11. Koelle DM, Huang ML, Chandran B, Vieira J, Piepkorn M, Corey L. Frequent detection of Kaposi’s sarcoma-associated herpesvirus (human herpesvirus 8) DNA in saliva of human immunodeficiency virus-infected men: clinical and immunologic correlates. J Infect Dis 1997; 176: 94–102.
12. Pellett PE, Spira TJ, Bagasra O, Boshoff C, Corey L, de Lellis L, et al. Multicenter comparison of PCR assays for detection of human herpesvirus 8 DNA in semen. J Clin Microbiol 1999; 37: 1298–1301.
13. Loiseau P, Mariotti M, Corbi C, Ravera N, Girot R, Thauvin M, et al. Prevalence of hepatitis G virus RNA in French blood donors and recipients. Transfusion 1997; 37: 645–650.
14. Biagini P, Gallian P, Touinssi M, Cantaloube JF, Zapitelli JP, de Lamballerie X, et al. High prevalence of TT virus infection in French blood donors revealed by the use of three PCR systems. Transfusion 2000; 40: 590–595.
15. Biagini P, Gallian P, Attoui H, Cantaloube JF, Touinssi M, de Micco P, et al. Comparison of systems performance for TT virus detection using PCR primer sets located in non-coding and coding regions of the viral genome. J Clin Virol 2001; 22: 91–99.
16. Biagini P, Gallian P, Cantaloube JF, Attoui H, de Micco P, de Lamballerie X. Distribution and Genetic Analysis of TTV and TTMV Major Phylogenetic Groups in French Blood Donors. J Med Virol 2006; 78: 298–304.
17. Yzebe D, Xueref S, Baratin D, Bouletreau A, Fabry J, Vanhems P. TT virus. A review of the literature. Panminerva Med 2002; 44: 167–177.
18. Ukita M, Okamoto H, Kato N, Miyakawa Y, Mayumi M. Excretion into bile of novel unenveloped DNA virus (TTV) associated with acute and chronic non-A to G hepatitis. J Infect Dis 1999; 179: 1245–1248.
19. Maggi F, Pifferi M, Fornai C, Andreoli E, Tempestini E, Vatteroni M, et al. TT virus in the nasal secretions of children with acute respiratory diseases: relations to viremia and disease severity. J Virol 2003; 77: 2418–2425.
20. Inami T, Konomi N, Arakawa Y, Abe K. High prevalence of TT virus DNA in human saliva and semen. J Clin Microbiol 2000; 38: 2407–2408.
21. Garbuglia AR, Iezzi T, Capobianchi MR, Pignoloni P, Pulsoni A, Sourdis J, et al. Detection of TT virus in lymph node biopsies of B-cell lymphoma and Hodgkin’s disease, and its association with EBV infection. Int J Immunopathol Pharmacol 2003; 16: 109–118.
22. Cacoub P, Rosenthal E, Gerolami V, Hausfater P, Ghillani P, Sterkers Y et al. Transfusion-associated TT virus co-infection in patients with hepatitis C virus is associated with type II mixed cryoglobulinemia but not with B-cell non-Hodgkin lymphoma. Clin Microbiol Infect 2003; 9: 39–44.
23. Zhong S, Yeo W, Tang M, Liu C, Lin XR, Ho WM, et al. Frequent detection of the replicative form of TT virus DNA in peripheral blood mononuclear cells and bone marrow cells in cancer patients. J Med Virol 2002; 66: 428–434.
24. de Villiers EM, Schmidt R, Delius H, Zur Hausen H. Heterogeneity of TT virus related sequences isolated from human tumour biopsy specimens. J Mol Med 2002; 80: 44–50.

