


Gábor Tax1, Edit Urbán2, Zsuzsanna Palotás3, Róbert Puskás4, Zoltán Kónya4,5, Tamás Bíró6, Lajos Kemény1,3# and Kornélia Szabó3#
1Department of Dermatology and Allergology, University of Szeged, 2Institute of Clinical Microbiology, University of Szeged, 3MTA-SZTE Dermatological Research Group, 4Department of Applied and Environmental Chemistry, University of Szeged, 5Reaction Kinetics and Surface Chemistry Research Group of the Hungarian Academy of Sciences, Szeged, 6DE-MTA “Lendület” Cellular Physiology Research Group, Departments of Physiology and Immunology, Faculty of Medicine, University of Debrecen, Debrecen, Hungary
#These two authors contributed equally to this work.
Propionibacterium acnes is an important member of the skin microbiome. The bacterium can initiate signalling events and changes in cellular properties in keratinocytes. The aim of this study was to analyse the effect of the bacterium on an immortalized human keratinocyte cell line. The results show that various P. acnes strains affect the cell-growth properties of these cells differentially, inducing cytotoxicity in a strain-specific and dose-dependent manner. We propose that bacterially secreted propionic acid may contribute to the cytotoxic effect. This acid has a role in maintaining skin pH and exhibits antimicrobial properties, but may also have deleterious effects when the local concentration rises due to excessive bacterial growth and metabolism. These results, together with available data from the literature, may provide insight into the dual role of P. acnes in healthy skin and during pathogenic conditions, as well as the key molecules involved in these functions. Key words: immortalized keratinocyte cell line (HPV-KER); Propionibacterium acnes; acne vulgaris; short-chain fatty acid; propionic acid.
Accepted May 27, 2015; Epub ahead of print Jun 3, 2015
Acta Derm Venereol 2015; XX: XX–XX.
Kornélia Szabó, MTA-SZTE Dermatological Research Group, Korányi fasor 6, HU-6720 Szeged, Hungary. E-mail: szabo.kornelia@med.u-szeged.hu
Human skin harbours a specialized microbiota. Although the existence of this microbial community has been known for a long time, its exact function and contribution to healthy skin functions, and its role in the initiation of skin diseases during pathogenic conditions, has not been clearly established (1, 2). A complex and dynamic interaction has been recognized between the microbes and skin cells, and an increasing volume of data indicates their importance for the development and maintenance of healthy skin (3). Under certain conditions that are currently not well defined, dysbiosis may lead to the pathogenesis of different skin diseases, including the most common and well-known multifactorial, chronic inflammatory skin disease, acne vulgaris (4).
According to our current understanding, acne lesions start to develop around puberty as a result of hormonal changes that include an androgen excess, leading to enhanced sebum secretion and the subsequent hypercolonization of the otherwise commensal P. acnes bacterium (5). In addition, abnormal keratinocyte and sebocyte function, as well as innate immune and inflammatory events, have been shown to play a major role during lesion formation (6). However, the exact cause and sequence of these events, and whether P. acnes plays role in the initiation of the unusual keratinocyte and sebocyte behaviour, are not known (7).
P. acnes exhibits cytotoxic and haemolytic activities (8) and can activate the innate immune system via different pathogen recognition receptors, such as the Toll-like receptors (TLR); TLR2 and TLR4 in particular have been implicated in these processes (9, 10). As a result of this activation, a transcriptional program is initiated that is responsible for the activation and subsequent nuclear translocation of the nuclear factor (NF)-kB transcription factor (11). These activities result in changes in the expression of several genes, including those for proinflammatory cytokines, chemokines and antimicrobial peptides, subsequently leading to innate immune and inflammatory events in the affected cells (9, 10).
Based on sequence differences in bacterial recA and tly genes and the results of multi-locus sequence typing (MLST) experiments, 6 phylogenetic groups within the P. acnes species have been described (IA1, IA2, IB, IC, II, III) (12, 13). It has been suggested that the strains belonging to these phylogenetic groups might exhibit different virulence properties as well as differentially affect the cellular and molecular properties of human keratinocytes (10, 14).
To systematically analyse this phenomenon, we investigated the effect of 3 selected P. acnes strains on various cellular responses of an in vitro cultured immortalized human keratinocyte cell line, HPV-KER (Polyánka & Szabó, in preparation). Results from different strains were compared in order to determine whether they exerted differential effects.
Our results show that the selected P. acnes strains have differential effects on the cellular properties (growth and viability) of HPV-KER cells. The observed effects are dose-dependent and correlate well with the bacterial load that is present in the environment of the human cells. Previous studies suggest that pattern-recognition receptor activation (e.g. mediated by TLR2 and TLR4) by conserved bacterial pathogen-associated molecular patterns and the subsequent initiation of the NF-kB-dependent transcriptional program may be responsible for the activation of an alternative keratinocyte program (10). Our results indicate that, apart from this signal transduction, strain-specific differences in the concentration of acidic metabolites secreted by P. acnes, including the short-chain fatty acid (SCFA) propionic acid (PA), may also contribute to the observed effects.
MATERIALS AND METHODS (See Appendix S11)
RESULTS
Strain-specific effects of P. acnes on cellular properties of HPV-KER cells
In order to systematically analyse the effect of different P. acnes strains on the cellular properties of human in vitro cultured immortalized keratinocytes a novel cell line, HPV-KER, was used. These were shown to grow in monolayer, exhibit keratinocyte-like morphologies and respond to the presence of P. acnes with similar gene expression changes to those of normal human adult keratinocytes (NHEK) cells (Fig. S11).
To monitor growth properties, HPV-KER cells were seeded in 96-well plates at different cell densities (5000, 10,000, 15,000 cells/well) and impedance-based analysis was performed using the xCELLigence system. Cell index (Ci) values were measured every 60 min for 72 h and plotted as a function of time (Fig. S21).
The cells started to attach to the surface of the wells within 3 h, indicated by a rapid increase in Ci values, and, after a short lag period, they entered a growth phase. Plates seeded with 15,000 cells/well reached confluence 36 h after plating, whereas plates seeded with other cell densities remained in the growth phase during the entire experiment. Plating densities of 10,000 cells/well were used in experiments to detect changes in HPV-KER proliferation in response to P. acnes treatment.
For determining the strain-specific effects of P. acnes treatment, the experiment was repeated with the addition of the P. acnes strains 24 h after plating (Fig. 1).
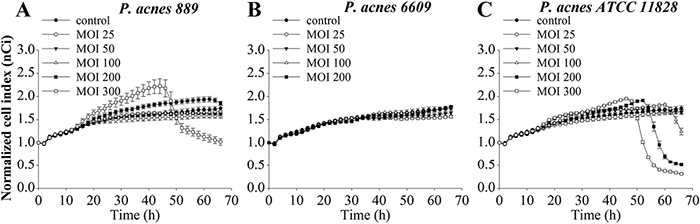
Our results suggest that, when using low multiplicity of infections (MOIs) (25 and 50), no P. acnes strain had a significant effect on the properties of HPV-KER cells compared with untreated controls. The cells continued to proliferate in the presence of the bacteria until they reached confluence, and subsequently entered a stationary phase, marked by steady normalized cell index (nCi) values. In contrast, when the P. acnes 889 strain was applied at a high dose (MOI=300), a rapid elevation of the nCi values was detected at approximately 15 h and continued until 45 h (Fig. 1A). However, when higher bacterial loads (MOIs of 200 or 300) of the 889 and ATCC 11828 strains were applied, a small increase followed by a sharp decrease were noted (Fig. 1A and C, respectively). Similar changes were not observed for treatment with P. acnes 6609 strain (Fig. 1B).
The above experiment was repeated, but this time instead of the HPV-KER cells, we treated NHEK ones using the same conditions. Our results indicate that NHEK cells responded to the presence of the P. acnes strains and exhibited nCi changes similar to those observed in the HPV-KER cells (Fig. S31).
P. acnes 889 and ATCC 11828 strains affect HPV-KER cell numbers
Ci changes can reflect differences in the number or the specific dimensions of cells attached to the electrodes. To determine the exact nature of the P. acnes-induced cellular events that corresponds to the observed nCi differences, we monitored the effect of the different P. acnes strains on the number of cells in the HPV-KER cell cultures.
High dose (MOI=300) of the P. acnes 889 strain resulted in increased cell numbers compared with untreated and low-dose treated samples (Fig. S4A1). None of the other treatments resulted in an increase in cell number. However, the rate of increase in the cell number decreased 12 h after high-dose treatment (MOI=300) with the ATCC 11828 strain.
High-dose treatment of the P. acnes 889 and ATCC 11828 strains induces microscopic changes in HPV-KER cells
To visualize cellular changes induced by P. acnes strains, we stained untreated and treated (MOI=300) HPV-KER cells with rhodamine-labelled phalloidin and performed a fluorescent microscopic analysis. We noted the presence of abnormally shaped rounded cells exhibiting irregular membrane morphology 48 h after treatment with the P. acnes 889 and ATCC 11828 strains (white arrows). This effect was not apparent in cells treated with the P. acnes 6609 strain (Fig. 2).
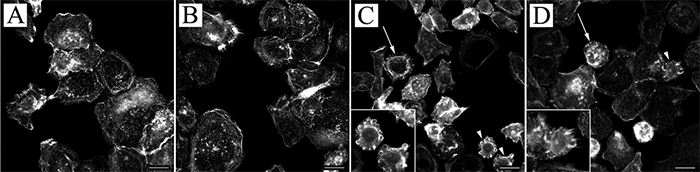
Fig. 2. Microscopic analysis of P. acnes-treated HPV-KER cells. Immortalized keratinocytes were stained with rhodamine-labelled phalloidin to visualize cytoskeletal F-actin bundles for an overall evaluation of cellular morphology. The presence of abnormally shaped, round cells was detected in the cultures treated with: (C) P. acnes 889 and (D) ATCC 11828 (multiplicity of infections 300), exhibiting a high degree of membrane irregularity (marked with white arrows and arrowheads), which was not apparent in: (A) control and (B) P. acnes 6609-treated cultures. (Arrowheads indicate cells depicted at high magnification in the lower left corner. Scale bars: 10 µm).
P. acnes-induced cytotoxicity is strain- and dose-dependent
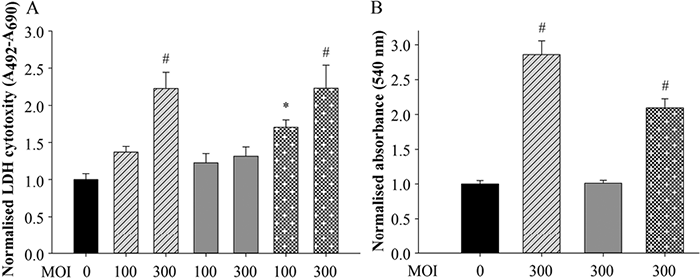
To determine whether P. acnes-induced cytotoxicity was due to damage of the keratinocyte membrane caused by the bacterium or by bacterially derived toxins, the amount of free lactate dehydrogenase (LDH) enzyme released to the supernatant of the damaged cells was determined using an LDH assay of keratinocytes treated with different doses (MOIs of 100 and 300) of the P. acnes strains.
Higher LDH levels were measured in cells treated with the 889 and ATCC 11828 strains. This effect appeared to be dose-dependent at 72 h post-treatment. In contrast, no differences were detected in LDH levels in cells treated with the P. acnes 6609 strain (Fig. 3A).
P. acnes exhibits a strain-specific haemolytic effect on human erythrocytes
To test whether the cytotoxic events caused by the selected P. acnes strains was specific to keratinocytes, we also treated washed human erythrocytes with the bacterium (MOI=300). Spectroscopic analysis was performed to quantify the amount of free haemoglobin released to the supernatant of the cell cultures as a result of possible membrane damage.
The free haemoglobin results were similar to the results of the LDH assay: treatment with the P. acnes 889 and ATCC 11828 strains increased the concentration of free haemoglobin in the culture supernatants at 72 h post-treatment, whereas treatment with the P. acnes 6609 strain had no such effect (Fig. 3B).
Some P. acnes strains increase the pH of HPV-KER cell cultures
To determine pH changes induced by P. acnes, HPV-KER cells were plated in serum-free Dulbecco’s Modified Eagle’s Medium, high glucose (DMEM-HG) media, which contains a pH-sensitive phenol-red dye. We observed enhanced acidification of cultures treated with the P. acnes 889 and ATCC 11828 strains by visual inspection. The extent of acidification was dose-dependent (Fig. S51).
P. acnes production of PA may contribute to media acidification and cellular changes in the HPV-KER cultures
The P. acnes bacterium generates SCFAs during metabolism. To determine whether the presence of SCFAs contributes to the observed cellular changes, we treated the HPV-KER cells with PA. Microscopic analysis of cells treated with SCFAs and rhodamine-labelled phalloidin revealed similar irregular membrane morphologies observed for P. acnes treatment. This result suggests that PA may be a factor leading to the observed keratinocyte morphological changes (Fig. 4).
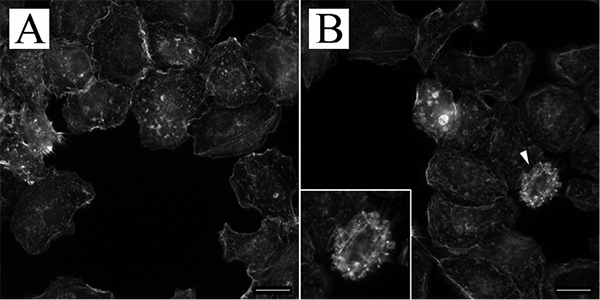
Fig. 4. Microscopic analysis of propionic acid (PA)-treated HPV-KER cells. (A) Control and (B) 48 h PA-treated HPV-KER cells were stained with rhodamine-labelled phalloidin and subjected to microscopic analysis. Cells exhibiting irregular membrane morphology were detected in the PA-treated cultures. (Arrowheads indicate cells depicted at high magnification in the lower left corner. Scale bars: 10 µm).
PA secretion of P. acnes is strain- and dose-dependent
To determine whether there are differences in the amount of P. acnes-secreted SCFAs, we treated HPV-KER cells with the bacterium (MOI 0 =100, 300), and the supernatant samples were subjected to mass spectrometry (MS) analysis.
The presence of acetic acid (AA) and PA were detected in all samples. While the AA levels were similar in all cases, the amount of PA varied in a strain-, and dose-dependent manner at 72 h post-treatment (Fig. 5); higher levels were detected when higher bacterial loads (MOI =300) of the 889 and ATCC 11828 strains were applied (Fig. 5D and H, respectively). Low levels of butyric acid (BA) were also detected in the high-dose (MOI = 300) P. acnes 889-treated HPV-KER culture supernatants (Fig. 5D).
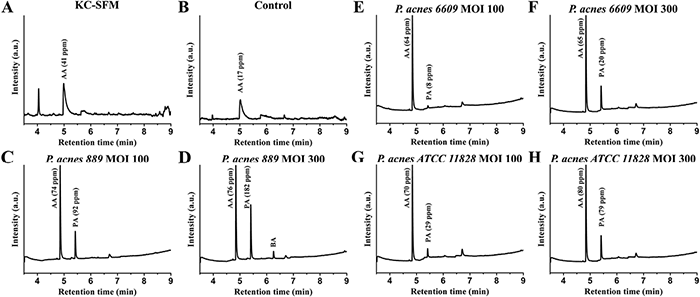
Fig. 5. Identification of short-chain fatty acids (SCFAs) in HPV-KER culture supernatant samples. (C–H) HPV-KER cells were treated with P. acnes strains. After 72 h, released SCFAs were measured by mass spectrometry (MS). While the level of acetic acid (AA) was relatively stable in the samples, the propionic acid (PA) levels exhibited strain- and dose-dependent differences; higher amounts detected in the high-dose (MOI=300) P. acnes 889 and ATCC 11828-treated cultures. In (A) the control medium (keratinocyte serum-free medium (KC-SFM), and (B) the untreated HPV-KER supernatant samples the presence of PA and butyric acid (BA) were not detected.
In order to prove that the detected SCFAs are bacterially-derived, we repeated the experiments, but this time in the absence of HPV-KER cells. While some AA was already present in the culturing media, bacterial fermentation clearly resulted in the release of additional amounts. PA and BA were only detected in the bacterial-treated samples under certain conditions (Fig. S61).
Combined treatment with the P. acnes 6609 strain and PA induces cytotoxicity
To determine whether PA plays a role in the bacterium-induced cytotoxic effects, we treated washed human erythrocytes with the P. acnes 6609 strain (MOI=300) in the presence of 1, 2 and 5 mM PA (Fig. S71). While this strain did not induce cytotoxicity alone, it led to the appearance of increasing levels of free haemoglobin parallel with the increasing concentrations of PA (Fig. S61).
DISCUSSION
Human skin harbours a specialized microbial flora that is located mostly on the surface of the skin and within the pilosebaceous unit (15). The exact function of this microbial community is not yet fully understood; however, it has been suggested that the interaction of the skin flora with skin cells is important for the maintenance and promotion of healthy skin functions (3). The composition of skin microbiota differs on various body parts, and although P. acnes is a major constituent, it is especially abundant in the sebum-rich regions (16). This Gram-positive Actinobacteria resides preferentially on the face, shoulders, upper chest and back, which are also the regions that are most affected by acne vulgaris during puberty. Thus, P. acnes has long been believed to play a role in the pathogenesis of this skin disease (4).
Detailed microbiological and, later, sequence-based phylogenetic analyses of various clinical isolates have indicated that this bacterial species is not homogenous, and different phylogenetically distinct subgroups (phylogroups) have been identified (13). It has been suggested that different strains belonging to these phylogroups may differentially affect various cellular and molecular properties of certain human cell types (10, 17).
In order to gain a better understanding of the properties of the interaction between the skin-colonizing P. acnes bacterium and the epidermal keratinocytes, we set up and systematically analysed an in vitro model system, using HPV-KER, a human immortalized keratinocyte cell line, previously established and characterized in our laboratory. We wanted to use a cell line that is easy to maintain, the availability of which is not limited, but which responds to the presence of the bacterium in vitro similarly to NHEK cells. In preliminary experiments we evaluated the well-known and widely used HaCaT cells. However, possibly because of their abnormally high NF-kB transcription factor levels (18), which is a known mediator of the TLR-mediated signalling events, HaCaT cells responded differently compared with NHEKs. In contrast, the newly established immortalized keratinocyte cell line HPV-KER reacted similarly to the presence of the bacterium (Figs S1–S31).
Next, we systematically analysed the effect of selected P. acnes strains (889, 6609 and ATCC 11828) representing different phylogroups on HPV-KER cells. Using a detailed, real-time, label-free, impedance-based approach, we found clear strain- and dose-specific differences.
High-dose treatment with P. acnes 889 leads to a transient increase in the number of HPV-KER cells up to 12 h after treatment. This presumed increase in proliferation is very intriguing, as one of the initial and characteristic cellular changes that take place in vivo during acne lesion development is ductal hypercornification. This hyperproliferation and abnormal differentiation of keratinocytes is particularly notable for keratinocytes lining the ductal region of the pilosebaceous units (19, 20). In the follicles, P. acnes mostly comes into direct contact with differentiated keratinocytes, whereas proliferating basal cells, that are modelled in our system reside in deeper tissue compartments. Our results suggest that selected P. acnes strains may contribute to these early changes in areas where the local presence of the bacterium reaches a certain threshold cell density through the presence of keratinocyte and/or bacterially-derived secreted factors that may reach these cells.
A drop in nCi values was noted in cultures treated with high doses of P. acnes 889 and ATCC 11828. Microscopic analysis of these cultures revealed the presence of morphologically distinct, shrunken cells exhibiting extensive membrane irregularities. Recently, it was shown that these types of membrane reorganizations often occur in response to cellular damage caused, for example, by mechanical cell injury or the presence of various pore-forming toxins. Injured cells form blebs to localize and delimit the damaged area in an attempt to limit the loss of cellular constituents and to allow repair (21). We hypothesize that the observed morphological changes and, in the case of severe, irreparable damage, cell morphological changes and death, are responsible for the observed drop in nCi values detected in our experiments. These events did not appear to be the result of a cell-type-specific interaction: similar effects were also detected in washed human erythrocytes and other cell types treated with P. acnes (8, 14).
The production of bacterium-derived secreted pore-forming exotoxins may be responsible for the observed changes. One such molecule, the P. acnes Christie, Atkins, Munch-Peterson (CAMP) factor, has been shown to be secreted at a different rates by various strains (8, 22). However, the observation that the cellular changes correlated with marked pH changes in the cultures led us to hypothesize that these effects are due to PA, a P. acnes metabolic product resulting from bacterial fermentation (23). This hypothesis is supported by reports that PA causes cytotoxicity in various cell types (14, 23). We chose to test this hypothesis with in vitro experiments. Our MS analysis revealed the presence of AA and PA in the supernatant of P. acnes-treated HPV-KER cells, and strain- and dose-dependent differences were measured in the PA levels. We also measured BA, but only in the high-dose P. acnes 889-treated cells. Next, we proved that the detected PA, BA and the majority of AA is of bacterial origin. Based on these results, we hypothesized that PA may be responsible for the previously observed effects, and we tested this idea in further in vitro experiments (Fig. S61).
Available data on the concentration of PA in the pilosebaceous unit are limited. However, measurements in other, more easily accessible, organs (e.g. the colon), where fermenting bacterial species are present suggests that PA levels can be as high as 20–140 mM (24). These concentrations are sufficient to control pathogenic microbes: minimal bactericidal or fungicidal PA concentrations for different species were found to be 10–25 mM (25). Based on these data, we chose PA concentrations of 1–10 mM for our in vitro experiments. Our results indicate that these treatments were sufficient to induce cellular changes in HPV-KER cells.
The function of PA in the skin is expected to be complex. PA contributes to skin acidification, which inhibits the colonization and growth of harmful invaders (26). In addition, PA exhibits antimicrobial properties that may be independent of its acidic nature (25). High PA concentrations, however, together with other bacterial factors, may also have deleterious effects, leading to cellular damage and, thus, compromising the integrity of the epidermal barrier.
Our observations suggest that the cellular events that are initiated by the bacterium are strongly dose-dependent. This challenges the generally accepted view that there is no correlation between the bacterial load and the presence of acne (27). However, our findings support novel studies suggesting that P. acnes is detected more frequently in acne vulgaris than in control skin samples (28). The differences between earlier and current results may reflect variations in the sampling techniques used. The bacterium is present in follicles as a biofilm, making quantitative studies more challenging (28, 29).
One of the studied strains, P. acnes 6609 appeared to be the least effective in our in vitro assays. MS analysis of the bacterial treated HPV-KER supernatants showed that the PA levels were the smallest in these samples. This appears to be the direct consequence of the impaired growth properties of P. acnes 6609, compared with the other 2 studied strains (Fig. S81). Such differences may contribute to the determination of individual acne lesion severity in the carriers.
Similar to other members of the skin microbiome, various P. acnes strains activate pattern-recognition receptors to initiate signalling events in the cytoplasm of the affected cells (10). Thus, the presence of the bacterium may be responsible for the initiation of an alert state in the surrounding cells within the follicles. According to our current model, a plug closes the ductal region of the pilosebaceous unit during comedo formation. The environment within the follicle (increasingly anaerobic conditions, constant temperature and moisture, continuously generated sebum) aids the growth of P. acnes. The plug prevents sebum and bacterially-derived metabolic products from being emptied from the follicles to the skin surface, generating a gradual change in the microenvironment. P. acnes is an anaerobic, aerotolerant species, and as a result has the ability to grow under both anaerobic and microaerobic conditions. Thus, in the healthy follicles it is capable of oxidative phosphorylation under microaerophilic conditions. During comedo formation, the increasingly anaerobic conditions may induce a switch in the bacterial metabolism to fermentation of the sebum components and the subsequent release of SCFAs (30). As a result, excess acidification and increased concentration of certain molecules, such as PA, may lead to keratinocyte cytotoxicity, compromising the comedo wall and making it more vulnerable. This vulnerability, together with the increasing concentrations of molecules that promote innate immune and inflammatory events, and the severity of lesions, increases. The effect of PA is strikingly similar in the pathogenesis of periodontitis, another bacterially-mediated human inflammatory disease (31).
Together with available data from the literature our results may enhance the understanding of the dual role of P. acnes in healthy skin and during pathogenic conditions.
ACKNOWLEDGEMENTS
The authors wish to thank Tanácsné Bajkán Andrea for her excellent technical assistance and Viharosné Dósa-Rácz Éva for the statistical analysis. This work was funded by The Hungarian Scientific Research Fund (OTKA NK105369) and the European Union and the State of Hungary, co-financed by the European Social Fund in the framework of TÁMOP–4.2.2.A-11/1/KONV-2012-0035 and TÁMOP-4.2.4.A/2-11/1-2012-0001 grants. Kornélia Szabó is a recipient of the János Bolyai Research Scholarship from the Hungarian Academy of Sciences. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
The authors have no conflicts of interest to declare.

1http://www.medicaljournals.se/acta/content/?doi=10.2340/00015555-2154
REFERENCES


