


1Department of Dermatology, Venereology and Allergology and 3Institute of Biochemistry I, Clinic of the Goethe University, Frankfurt, Germany, and 2Department of Dermatology, Medical University of Graz, Graz, Austria
#These authors are both first authors and contributed equally to this work.
The mTOR (mechanistic target of rapamycin) inhibitor rapamycin has long been known for its immune suppressive properties, but it has shown limited
therapeutic success when given systemically to patients with psoriasis. Recent data have shown that the mTOR pathway is hyperactivated in lesional psoriatic skin, which probably contributes to the disease by interfering with maturation of keratinocytes. This study investigated the effect of topical rapamycin treatment in an imiquimod-induced psoriatic mouse model. The disease was less severe if the mice had received rapamycin treatment. Immunohistological analysis revealed that rapamycin not only prevented the activation of mTOR signalling (P-mTOR and P-S6 levels), but almost normalized the expression of epidermal differentiation markers. In addition, the influx of innate immune cells into the draining lymph nodes was partially reduced by rapamycin treatment. These data emphasize the role of mTOR signalling in the pathogenesis of psoriasis, and support the investigation of topical mTOR inhibition as a novel anti-psoriatic strategy.
Key words: psoriasis; imiquimod; rapamycin; mTORC.
Accepted Jun 8, 2017; Epub ahead of print Jun 9, 2017
Acta Derm Venereol 2017; 97: xx–xx.
Corr: Claudia Buerger, Department of Dermatology, Venereology and Allergy, Clinic of Goethe-University, Theodor-Stern-Kai 7, DE-60590 Frankfurt am Main, Germany. E-mail: Claudia.Buerger@kgu.de and Peter Wolf, Department of Dermatology, Medical University of Graz, Auenbruggerplatz 8, AT-8036 Graz, Austria. E-mail: peter.wolf@medunigraz.at
Psoriasis is one of the most common inflammatory skin disorders, affecting approximately 3% of the world’s population (1). Several cellular changes in the skin, such as epidermal hyperplasia (acanthosis) and aberrant differentiation (parakeratosis), along with dermal inflammatory infiltrates related to innate and adaptive immune response and increased tortuous capillaries (angiogenesis) define its severity. This series of events leads to formation of sharply demarcated, red scaly
erythematous plaques on the skin, which can be itchy and painful (2). Psoriasis is heterogeneous in nature and exists in several distinct forms, in which approximately 80% of cases are classified as mild (defined by a Psoriasis Area Severity Index (PASI) ≤ 10) and in which the extent of body surface area covered varies widely from patient to patient (1). The severity of the disease usually determines the therapeutic approach to controlling psoriasis. Approximately 70–80% of all patients with psoriasis who have mild disease can be treated adequately with topical therapies, using agents such as corticosteroids, vitamin D analogues, topical retinoids and calcineurin inhibitors. For moderate-to-severe psoriasis (e.g. affecting large surface areas) a well-established treatment regimen is a combination of topical agents and phototherapy or systemic drugs (3, 4). In recent years no novel substances have been developed for topical treatment, and newly introduced agents have mainly been analogues, derivatives or new formulations of already known agents (5).
PI3-kinase (PI3-K) and its downstream effector, mTORC1, being fundamental components of immune cell-signalling networks, play a crucial role in skin homeostasis and morphogenesis, especially in the regulation of keratinocyte differentiation and epidermal stratification (6–8). We have shown previously that the mTOR kinase itself and its downstream target, ribosomal protein S6, are hyper-activated in psoriatic lesions (9, 10). In addition, the PI3-K/mTOR pathway is thought to play a role in Th1-Th2-Th17 imbalance in the pathogenesis of psoriasis (11). The mTOR inhibitor rapamycin, also known as sirolimus, was initially investigated as an antifungal agent and was used for its immunosuppressant properties (12) and also showed anti-tumourigenic potential. Rapamycin binds to the cytoplasmic protein FKPB-12, which interacts with mTORC1, thus blocking the mTOR target proteins S6 and 4E-BP that are essential for protein synthesis and thereby for cell growth and proliferation (13). The aberrant activation of mTOR in different cancers and in epidermal tumours has promoted the investigation of rapamycin as an anti-tumourigenic therapy and enabled further development of a range of small molecule inhibitors that may prove effective in disease control (14).
There have not been many large-scale trials to validate the potential of rapamycin as an effective topical anti-inflammatory agent in psoriasis (15), and few studies have tested its anti-psoriatic effect after oral administration (16–18). However, topical rapamycin treatment for tuberous sclerosis complex (TSC) is well documented. In TSC, genetic mutations of the TSC1 or TSC2 gene inhibit the association of their protein products, tuberin and hamartin, leading to an aberrant upregulation of the mTOR signalling pathway. Reportedly, topical rapamycin showed remarkable efficacy in treating angiofibromas of patients with TSC (19). Thus, to investigate the effectiveness of topical rapamycin treatment and the role of mTOR in psoriasis development, we used a murine imiquimod (IMQ) model. IMQ, a Toll-like receptor (TLR)7/8 agonist, can induce de novo and exacerbate pre-existing psoriasis lesions, which is critically dependent on the interleukin (IL)-23/IL-17 axis and IL-22. Histologically, IMQ treatment results in keratinocyte hyperproliferation, causing abnormal epidermis differentiation with increased angiogenesis, and high infiltration of immune cells (20, 21). Studies involving the use of this mouse model have shown activation of the PI3K-mTOR cascade similar to human psoriasis (22), and blockade of PI3-K in this models improved clinical symptoms (23). These findings demonstrate the functional importance of the mTOR pathway in the pathogenesis of psoriasis, and strongly advocate further development and investigation of topical rapamycin as an anti-psoriatic strategy.
The aim of this study was to investigate the efficacy of topical rapamycin, using the IMQ-induced psoriatic phenotype in mice, specifically regarding disease progression and skin barrier development, as well as its effect on immune cells, with the ultimate goal of identifying novel anti-psoriatic treatment options.
Aldara® (IMQ) 5% cream (MEDA Pharmaceuticals, Vienna, Austria) was used for inducing psoriatic skin changes in the mice. 1% Rapamycin (Merck Calbiochem, Darmstadt, Germany) ointment was made by mixing 50 mg petroleum jelly with 10 μl of the corresponding rapamycin stock, 50 mg/ml. P-mTOR S2448 #2976 and P-S6 S235/6 #2211 antibodies were from Cell Signaling Technology, Frankfurt, Germany. Keratin 6 antibody (Ks6.KA12) from Thermo Scientific, Darmstadt, Germany and caspase-14 antibody (NB100-56126) was from Novus Biologicals, Wiesbaden, Germany. NIMP-R14 Ly-6G/C (ab2557) was from Abcam, Cambridge, UK. Fluorescein isothiocyanate (FITC)-conjugated anti-CD3, phycoerythrin (PE)-conjugated anti-CD4, PE-conjugated anti-CD11c, allophycocyanin (APC)-conjugated anti-CD11b, FITC-conjugated anti-F4/80, APC-conjugated anti-B220, PE-conjugated anti-langerin, APC-conjugated anti-EpCAM all were from eBioscience, Darmstadt, Germany, while peridinin chlorophyll protein complex (PerCP/Cy5)-conjugated anti-Siglec-H was from Biolegend, Koblenz, Germany and PerCP-conjugated anti-CD8 was purchased from BD Bioscience, Heidelberg Germany.
BALB/c mice were purchased from Charles-River, Sulzfed, Germany and housed in the animal facility of the Center for Medical Research, Medical University of Graz, Austria. All procedures to which the mice were subjected were approved by the Austrian Government, Federal Ministry for Science and Research, through protocol number BMWF-66-010/0032-11/3b/2013. Female mice at the age of 7–8 weeks were shaved on the back skin 48 h before the start of the study. Experimental groups included 3 regimes: vehicle + no IMQ (control), vehicle + IMQ (IMQ) and 1% rapamycin + IMQ (RAPA+IMQ). Application of 50 mg freshly made 1% rapamycin ointment or vehicle (petroleum jelly) was made to the dorsal skin using a flat spatula in the morning. Six hours later 62.5 mg IMQ cream was applied. Both rapamycin and IMQ treatments were given for 4 days, while rapamycin alone was applied for 1 additional day.
To measure the severity of inflammation on the back, a scoring system similar to the human Psoriasis Area and Severity Index (PASI) score was used, as described previously (24). In brief, ery-thema, infiltration and scaling of the skin were scored “blindly” on a scale from 0 to 3, as follows: 0: none; 1: slight; 2: moderate; 3: severe. The single scores were summed, resulting in a theoretical maximal total score of 9. In addition, skin thickness was assessed by measuring the double skin-fold thickness (DSFT) of dorsal skin of the mice with a spring-loaded engineer’s micrometer (Mitutoyo, Japan) throughout the experiment. Percentage reduction in specific skin swelling (S) was determined using the following formula: (1-(SRapa+IMQ–Scontrol/SIMQ–Scontrol))×100. Mice were euthanized with an overdose of isoflurane and all efforts were made to minimize suffering. Mice were sacrificed 24 h after the last application of rapamycin on day 6 and blood, skin and lymph nodes samples were collected for further analysis. Total and differential blood count was determined using a MS9-5 haematology analyser from Melet Schloesing.
For quantification of vascularization, the dorsal skin and adjacent soft tissue of the mice was prepared for taking photographs from the reverse site and the number of vascular bifurcations per dorsal skin sample was counted, as described previously (25).
Skin samples were fixed in 4% paraformaldehyde, paraffin embedded and stained with haematoxylin and eosin (H&E). The thickness of epidermis was measured and number of layers was measured in 5 microscopic fields at 20× magnification per mouse. The results were expressed as mean values. For immunohistochemistry (IHC), paraffin sections were processed routinely. Primary antibody was applied overnight after antigen retrieval with citrate solution pH 6. Histofine Simple Stain AP Multi (Medac Diagnostika) was used for detection, according to the manufacturer’s instructions. Images were acquired with a Nikon Eclipse slide scanning microscope.
The Opal™ 6-Color Fluorescent IHC Kit (Perkin Elmer) was used according to the manufacturer’s instructions. Slides were stained with primary antibodies targeting Ki-67, Keratin 10 (both from Abcam), Keratin 14, Loricrin and Involucrin (Biolegend). The Vectra® 3 automated quantitative pathology imaging system (Perkin-Elmer) was used for image acquisition and InForm software was used to score the relative abundance of cells expressing a certain marker protein in the epidermis or dermis upon tissue segmentation.
Isolated lymph nodes were filtered through a sterile sieve (40 μm) to obtain single-cell suspension. Cells were washed twice, and 1.5×107 cells were stained for 20 min in the dark at room temperature (RT) with fluorescent-labelled antibodies diluted in phosphate-buffered saline (PBS)/0.3% bovine serum albumin (BSA). Stained cells were washed with washing buffer and resuspended in 300 µl PBS with 30 µl Sytox (Life Technologies, USA). Labelled cells were quantified on a flow cytometer (FACScan or FACSCalibur; BD Biosciences) and analysed using CellQuest software (BD Biosciences). The viability of the cells was checked by staining with Sytox.
All macroscopic readings and scorings of microscopic slides were conducted in a blinded fashion. Data are presented either as a bar graph of the mean ± standard error of the mean (SEM) or as scatter blot of individual data-points with mean ±SEM. Statistical differences among experimental groups were determined using analysis of variance (ANOVA) and post-hoc Bonferroni multiple analysis or Mann–Whitney U test after testing for normality, whatever appropriate, using GraphPad Prism. The statistical test chosen for each experiment is indicated in the figure legend. Statistical significance was set at p ≤ 0.05 and levels of significance were given as *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001 and ****p ≤ 0.0001.
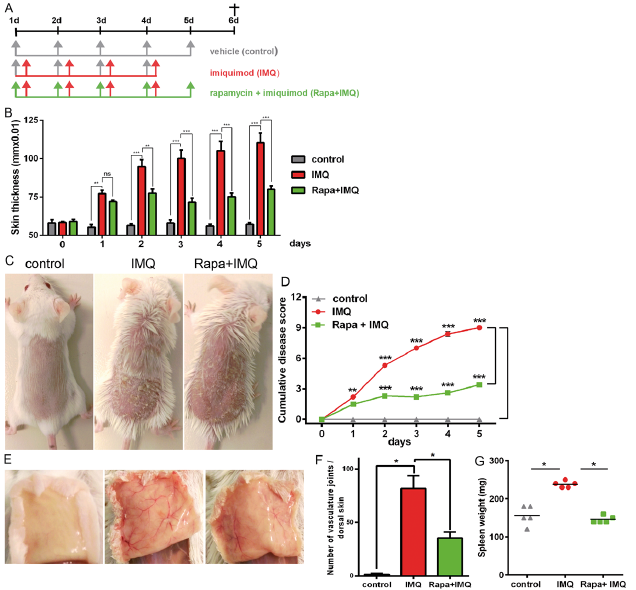
The efficacy of topical rapamycin treatment in IMQ-induced psoriasis-like skin lesions was evaluated in the mouse model, as shown in the study plan (Fig. 1A). Rapamycin cream or vehicle was applied in the morning for 5 days, followed by IMQ cream, after a 6 h gap, on the shaved back skin of BALB/c mice for the first 4 consecutive days. Two days after the start of IMQ application, the back skin of the mice started to display signs of erythema and swelling. In comparison with the IMQ-only treated group, the rapamycin-treated group showed statistically significantly reduced specific skin swelling of 24% from day 2 and this difference was further increased to 49% on day 3 (p ≤ 0.01), reaching a peak difference of 64% at day 4 and 61% at day 5 (p ≤ 0.001) (Fig. 1B). The characteristic features of IMQ-induced skin inflammation (erythema, infiltration and scaling) was scored daily throughout the experiment, and the single (Fig. 2) as well as the total scores showed significant reduction with topical rapamycin compared with the IMQ-only group (Fig. 1 C, D). While IMQ-treated mice showed severe induction of neovascularization in the dorsal skin compared with the control group, rapamycin prominently reduced neovascularization (Fig 1E, F). We consistently found significant enlargement of the spleen through induction of inflammation by IMQ, which could be rescued by rapamycin treatment (Fig. 1G). In summary, rapamycin treatment resulted in significant improvement in the psoriasis-like skin phenotype induced by IMQ.
Fig. 1. mTOR (mechanistic target of rapamycin) inhibition ameliorates skin and systemic symptoms of imiquimod-induced psoriasis. (A) Study plan: BALB/c mice were treated on the dorsal skin for 4–5 days by 3 different treatment regimes, as described in the materials and methods section. (B) Double skin-fold thickness (DSFT) assessment of dorsal skin of the mice measured throughout the experiment. (C) Macroscopic presentation of mice from the 3 treatment regimes. (D) Cumulative disease severity score during the course of the experiment. (E) Reduced neoangioenesis formation observed in dorsal skin of rapamycin-treated mice compared with imiquimod (IMQ)-treated ones. (F) Macroscopic bifurcations were counted as a measure of blood vessel formation. (G) Spleens were prepared from each mouse and weighed. All data shown is from 1 representative experiment, with n=5 per treatment group and error bars representing standard error of the mean (SEM). Statistical differences were determined by using 2-way analysis of variance (ANOVA) (B, D) or Mann–Whitney U test (F, G) (*p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001).
Fig. 2. Detailed clinical disease score. The severity of inflammation on the back was assessed with a scoring system similar to the human Psoriasis Area and Severity Index (PASI) score. Erythema, infiltration and scaling of the skin were scored “blindly” on a scale from 0 to 3, as follows: 0: none; 1: slight; 2: moderate; 3: severe. Statistical differences were determined by 2-way analysis of variance (ANOVA) and Bonferroni multiple comparison (**p ≤ 0.01; ***p ≤ 0.001).
Analysis of haematoxylin and eosin (H&E)-stained sections showed strong epidermal thickening, hyperkeratosis and elongated rete ridges in IMQ-treated mice (Fig. 3A), while rapamycin treatment led to a significant reduction in epidermal thickness and keratinocyte layers, along with a complete decrease in neutrophil-rich micro-abscess formation (Fig. 3A–D).
Fig. 3. Rapamycin reduces inflamma-tion induced by dermal mTOR (mechanistic target of rapamycin) signalling. (A) Haematoxylin and eosin (H&E) staining of representative image from each group. Bars represent 100 μm. (B, C) Evaluation of histological features, including epidermal thickness and epidermal layers. (D) Immuno-histochemistry (IHC) staining for neutrophil-rich micro-abscesses with a Ly-6G/C antibody from 1 representative mouse from each group. Scale bars represent 20 μm. The number of neutrophil rich micro abscesses was counted in the whole epidermis sheet. Statistical differences were determined by Mann–Whitney U test (*p ≤ 0.05). (E–P) IHC staining with the indicated antibodies is shown from 1 representative mouse from each group. Scale bars represent 20 μm.
To check whether rapamycin was able to interfere with inflammation-dependent mTOR signalling in this model, the epidermis was stained for the activation of mTOR kinase, by namely phosphorylation of S2448 and activation of its downstream target, the ribosomal protein S6. Hardly any phosphorylation was detected in the control group (Fig. 3E), but IMQ treatment led to an increase in P-mTOR at S2448 (Fig. 3F), especially in the basal layer, as it can be also seen in human psoriasis. Rapamycin treatment strongly diminished mTOR activation (Fig. 3G). A similar pattern was seen for the ribosomal protein S6, with strong phosphorylation in supra-basal layers after the induction of dermal inflammation (Fig. 3I), which was clearly decreased through rapamycin treatment (Fig. 3J). Keratin 6 upregulation is a characteristic feature of hyperproliferative cells in supra-basal layers of psoriatic plaque. In our mouse model we found that keratin 6 expression was induced in supra-basal layers after the induction of inflammation (Fig. 3L) and was again reduced if mTOR was inhibited by topical rapamycin (Fig. 3M).
Caspase-14 is a crucial component of an intact skin barrier, contributing to terminal differentiation by processing filaggrin (26) and is reduced in psoriatic skin (27). We found that, after IMQ treatment, the expression of caspase-14 was reduced, which could be rescued by rapamycin treatment (Fig. 3N–P), arguing again for normalization of the skin phenotype after rapamycin treatment.
Furthermore, in order to analyse the impact of aberrant mTOR signalling on epidermal maturation and skin inflammation, we used a multiplex IHC staining system. Keratinocytes in the epidermis were identified using keratin 14 as a marker and the number of maturing keratinocytes was quantified by co-staining with markers for the different epidermal layers (Fig. 4A). While IMQ increased the number of proliferating cells, rapamycin clearly reduced the number of Ki-67-positive cells to normal levels (Fig. 4B). Keratin 10 as a marker of early differentiation was significantly reduced, from 33% to 13% and 10% of positive cells respectively, under inflammatory conditions. Interestingly, the reduced expression of keratin 10 could not be rescued by rapamycin treatment (Fig. 4C). However, IMQ treatment doubled the number of involucrin-positive cells, which was significantly repressed when rapamycin was added (Fig. 4D). Loricrin, as a marker of late expression, was reduced by approximately 50% (from 9% to 4% of positive cells) under IMQ-induced inflammation, and showed a small, but not significant, increase after mTOR inhibition (Fig. 4E). In addition, the number of immune cells in the epidermis and dermis was quantified using the same multiplex IHC approach, looking for CD3-, MHCII- and Ly6B-positive cells. There was a trend for a reduction in immune cells in the skin of rapamycin-treated mice compared with IMQ and control mice, but statistical significance was not reached (data not shown).
Fig. 4. Rapamycin treatment normalizes the epidermal differentiation pattern and extravasation of immune cells. (A) Multiplex immunohistochemistry (IHC) staining with antibodies specific for Ki-67, keratin 14 (K14), keratin 10 (K10), involucrin (IVL) and loricrin (LOR) was performed. Composite images from 1 representative mouse of each group are shown. Insert present close-up images from the same slides. (B–E) The number of positive cells for the specific marker (Ki-67, K10, IVL or LOR) of all (K14-positive) keratinocytes per mouse sample was quantified in %. Data are mean values ± standard error of the mean (SEM) from 1 experiment, with n=5 per treatment group. Statistical differences were determined by using 1-way analysis of variance (ANOVA) and Bonferroni multiple comparison (**p ≤ 0.01, ****p ≤ 0.0001).
To assess the systemic effects of topical rapamycin treatment, the number of cells of innate and adaptive immunity immigrated into lymph nodes was monitored using flow cytometry (Table I). Many more macrophages (F4/80+ CD11b+), dendritic cells (DCs) (CD11c+ CD11b+), plasmacytoid DCs (pDCs) (CD11c+ 120G8+ SiglecH+) and Langerhans cells (CD207+ EpCAM+) were seen in the lymph nodes of IMQ-treated mice compared with untreated controls. There was a trend of reduction for all those cells in lymph nodes with rapamycin treatment, which was most prominent for pDCs (Table I). The blood count at the end of the experiment was also monitored (Fig. 5) and it was observed that IMQ increased the absolute leukocyte number (mainly due to increased levels of monocytes and neutrophils), as it is well documented that IMQ treatment causes a shift from lymphoid to myeloid cells (20). Treatment with rapamycin normalized the absolute leukocyte count in IMQ-treated mice to a level similar to that of the control group (Fig. 5). However, we did observe a trend for a decrease in the absolute number of lymphocytes, which is a known possible transient side-effect, paired with the therapeutic effect of rapamycin (28). Interestingly, normalization of the IMQ-increased absolute numbers of neutrophils and monocytes was found with rapamycin treatment. This is consistent with the fact that the mTOR complex regulates and influences many pivotal immunomodulatory functions of innate immune cells (29). Collectively, our data indicate the effectiveness of topical rapamycin in ameliorating the IMQ-induced psoriatic phenotype.
Table I. Topical rapamycin reverse the shift from lymphoid to myeloid cells in imiquimod model of psoriasis
Fig. 5. Systemic effect of topical rapamycin on total and differential leukocyte count. Total leukocytes were counted per 103/µl from n=5 mice per treatment group and the percentages of monocytes, neutrophils and lymphocytes were determined. Statistical differences were determined by using Mann–Whitney U test (*p ≤ 0.05, **p ≤ 0.01).
The pathogenesis of psoriasis is complex and adequate models have to be used to investigate novel treatment options. Recent studies using RNA microarray analysis have confirmed that the disease signature observed in the IMQ mouse model reflects the human setting, specifically with respect to genes involved in epidermal development, validating the use of this model (30). Using this model, we observed that IMQ induced an epidermal signalling pattern of the mTOR pathway that resembled the situation in human psoriasis (9), which supported previous findings (22) and advocated again for the IMQ model. Topical rapamycin treatment ameliorated the severity of erythema and scaling in the psoriasis-like condition, paired with a reduction in skin thickness and a marked reduction in neovascularization in the skin.
Rapamycin also had a strong effect in the epidermis on activation of the mTOR pathway itself and expression of the putative effector protein keratin 6. Keratin 6 is upregulated in hyperproliferative cells in the suprabasal layers of psoriatic plaques (31) and its mRNA shows putative 5´TOP elements, which are specifically regulated by mTORC1. In addition, its expression is sensitive to rapamycin (32). Thus, inhibition of mTOR might reduce the aberrant expression of keratin 6 and thereby contribute to the normalization of keratinocyte maturation. We also found normalization of caspase-14 expression after rapamycin treatment, which is supported by a study showing that anti-inflammatory therapy restores epidermal caspase-14 expression (33). To investigate the effects of rapamycin on the epidermal barrier we used a quantitative multiplex immunostaining approach that revealed reduced expression of keratin 10 and loricrin and an increase in expression with aberrant distribution of involucrin by IMQ, which resembles the situation in human psoriasis (34). Rapamycin was able to restore these disturbed expression patterns for most of the markers, similar to what can be seen in human skin biopsies after treatment with biologics (35). Thus, rapamycin seems to be a differentiation-promoting agent, as was also observed for other natural compounds interfering with mTOR activity (36). In addition, rapamycin normalized the IMQ-dependent increase in Ki-67-positive keratinocytes, possibly due to a direct effect of rapamycin on proliferative control, underlining the role of mTORC1 in keratinocyte stem cell homeostasis (37). On the other hand, the decline in proliferative keratinocytes might be a consequence of the switch towards keratinocyte differentiation induced by mTOR inhibition.
mTOR is a critical regulator of immune function, and rapamycin has been widely used as an immunosuppressant, which was originally attributed to the role of mTOR in controlling immune cell proliferation. However, it is now clear that mTOR mediates divergent effects in immune cells (reviewed in (38)): mTOR is not only involved in Th1 and Th17 differentiation, but also controls leukocyte trafficking by determining the repertoire of adhesion and chemokine receptors. In particular, we observed a trend towards normalization of the flux of DCs from the skin into the lymph nodes. These cells express TLR7 endosomally and mTOR plays an important role for TLR-mediated activation. The normalized dermal vessel density resulting in a decreased flow of leucocytes to the skin might also contribute to the reduced influx of APCs from the skin into secondary lymphoid organs in rapamycin treated animals. Interestingly, vascular endothelial growth factor (VEGF), which is known to promote vascular permeability leading to enhanced leucocyte traffic into the skin, is also targeted by rapamycin (39). Through these combined mechanism topical rapamycin seems to normalize blood count parameters, flux into lymph nodes, spleen weight, and possibly also the number of immune cells in the skin.
Thus, from the data presented we cannot conclude whether keratinocytes or immune cells are the primary target of topical rapamycin. However, as we showed previously that PI-3K/mTOR signalling shows massive hyperactivation mainly in keratinocytes (9, 40), we assume that the major effect is due to the control of keratinocyte malfunction. In addition, a direct, but smaller, effect on cells of the adaptive and innate immune system is conceivable, as well as a secondary effect through reduced secretion of cytokines by keratinocytes.
Systemic mTOR therapy has shown promising outcome in autoimmune diseases, but was associated with significant side-effects. There are little data on the efficacy of topical rapamycin and its direct effect on skin structure and barrier integrity. However, in recent clinical studies topical rapamycin ointment not only showed adequate skin penetration along with a good safety profile, but also significantly ameliorated skin angiofibromas, which are characterized by hyperactivation of mTOR signalling (19). These outcomes, together with our findings, support the rationale that topical rapamycin is effective in the treatment of skin diseases and that topical rapamycin should be further evaluated, both alone for mild forms of psoriasis and in combination with systemic drugs for moderate to severe forms of the disease.
This work was funded by a grant to C.B. from the German Research Foundation (DFG) BU 1840/5-1, Germany and FWF Austrian Science Fund number W1241 to P.W.. N.S. was supported by the PhD program Molecular Fundamentals of Inflammation (MOLIN) from the Medical University of Graz, Austria.
The authors declare no conflicts of interest.


 Click to show fullsize
Click to show fullsize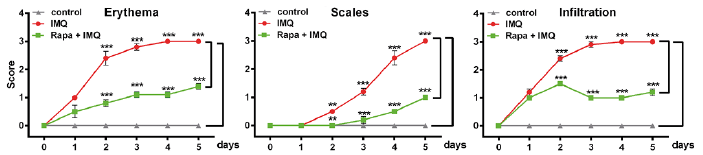 Click to show fullsize
Click to show fullsize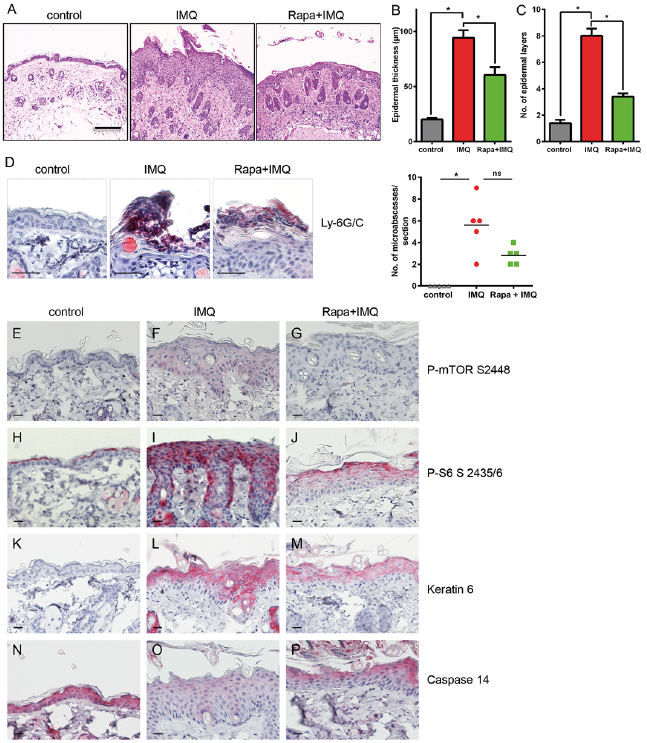 Click to show fullsize
Click to show fullsize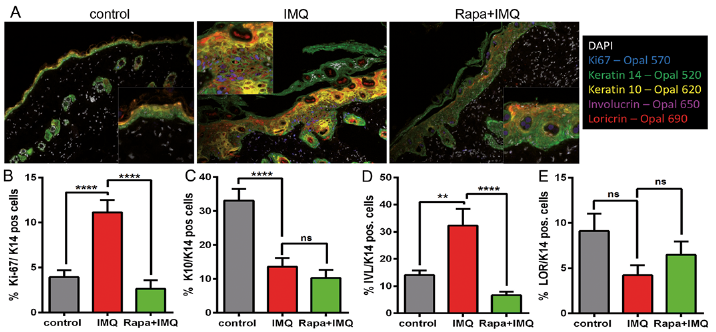 Click to show fullsize
Click to show fullsize Click to show fullsize
Click to show fullsize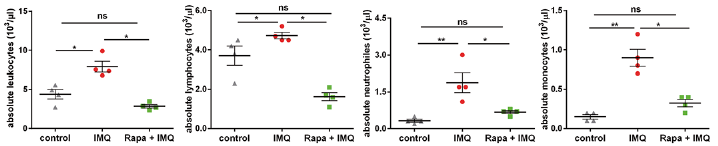 Click to show fullsize
Click to show fullsize