


1Division of Dermatology and Venereology, 2Division of Medical Diagnostics, Region Jönköping County, Jönköping, and 3Division of Cell Biology, Department of Clinical and Experimental Medicine, Linköping University, Linköping, Sweden
#These authors contributed equally to this work and are shared last author.
Changes in the skin microbiome have been shown to promote cutaneous inflammation. The skin microbiome of patients with chronic plaque type psoriasis was analysed before and after treatment with narrowband ultraviolet B (UVB). Swab samples of the microbiome were taken from lesional and non-lesional skin of 26 patients. Microbiotas were characterized by sequencing 16S rRNA bacterial genes on the Illumina MiSeq platform. Lesional skin microbiome diversity correlated with psoriasis severity (measured with the Psoriasis Area and Severity Index; PASI). There was a significantly lower abundance of the phylum Firmicutes and the genus Staphylococcus in lesional skin compared with non-lesional skin before UVB treatment. Responders (> 75% target Psoriasis Severity Index (PSI) improvement) had significantly lower abundance of the phyla Firmicutes in lesional and non-lesional skin and lower abundance of the genera Staphylococcus, Finegoldia, Anaerococcus, Peptoniphilus, Gardnerella, Prevotella and Clostridium in lesional skin after UVB treatment. Pseudomonas significantly decreased in lesional and non-lesional skin of treatment responders. These results suggest that skin microbiome alterations after UVB treatment could be related to treatment and treatment response.
Key words: psoriasis; microbiome; UVB treatment.
Accepted Nov 29, 2017; Epub ahead of print Dec 4, 2017
Acta Derm Venereol 2018; 98: XX–XX.
Corr: Oliver Seifert, Division of Cell Biology, Department of Clinical and Experimental Medicine, Linköping University, SE-581 83 Linköping, Sweden. E-mail: oliver.seifert@liu.se
Psoriasis is a chronic immune-mediated inflammatory condition with an estimated prevalence of 2–3% (1, 2). It is associated with a significantly increased risk for severe comorbidities, such as myocardial infarction and inflammatory bowel disease (IBD) (3–7). The immune response in psoriasis is characterized by hyperproliferation of keratinocytes, increased angiogenesis, and dermal infiltration of T cells, neutrophils and macrophages (8). Th17 T cells play a key role in psoriasis by stimulating dendritic cells and keratinocytes, leading to upregulation of antimicrobial peptides, inflammatory cytokines, and chemokines (9). In addition to similarities in the immune response pattern, IBD, such as Crohn’s disease, and psoriasis share multiple genetic susceptibility loci, e.g. IL23R, IL12B and TYK2 (10).
Genetic predisposition does not wholly explain the pathogenesis of psoriasis. It is also provoked by a variety of different environmental factors; for example, bacterial colonization in various parts of the body (1, 11, 12). The most well-known bacterial trigger is Streptococcal infections of the upper respiratory tract, which has long been recognized as a initiator of mainly guttate psoriasis (8). Other microorganisms commonly found on the skin, such as Staphylococcus aureus, Malassezia and Candida, have been linked with exacerbation of psoriasis (12–14), where the severity of psoriasis has been shown to correlate with enterotoxin production by isolated S. aureus strains (14).
Studies have shown that the intestinal microbiome of patients with IBD, such as Crohn’s disease, differs significantly from the microbiome of healthy individuals (15). A dysfunctional immune response to an altered intestinal microbiome plays a fundamental role in the pathogenesis of IBD in individuals with a certain genetic predisposition (16). Because of the similarities between Crohn’s disease and psoriasis, it has been proposed that psoriasis is caused by a breakdown of immune tolerance to the skin microbiota, where specific bacteria in the skin of genetically predisposed individuals activate the innate immune system, and, as a result, induce an adaptive immune response (17). If the specific microbiome pattern that initiates the psoriatic pathophysiology was found, it might be possible to treat the disease by “normalizing” the microbiome (18). The anti-inflammatory effect of “normalizing” the microbiome has been shown for atopic dermatitis (19, 20).
Only a few studies have investigated the skin microbiome in psoriasis, with inconsistent results (18, 21–23). One study suggests that the microbiome of psoriasis plaques is more diverse than unaffected skin sites (21), while another study found a trend towards decreased diversity of psoriatic lesions (18). A third study analysing the microbiome in skin biopsies, found no differences in alpha or beta diversity between lesional and non-lesional skin (22). Differences in Firmicutes and Actinobacteria found in one study (21) were not present in other studies when taking body sites into account and studying samples from trunk and limbs (22). Proteobacteria were found to be significantly more abundant in trunk psoriasis samples compared with a control group, but with no significant differences when comparing legs or arms (22). In 2 studies, the ratio of Streptococcus to Propionibacterium was higher in psoriasis compared with controls (21, 22). One study found no differences in the relative abundance of Firmicutes or Actinobacteria, but lesional skin displayed the highest intragroup diversity, while unaffected skin from patients with psoriasis was more similar to control skin (18). Recently, high-resolution shotgun metagenomics was used to characterize the microbiome of lesional and non-lesional skin. They found psoriatic ear sites to have decreased diversity, and associated lesional skin with an increased abundance of Staphylococcus, but no specific biomarker indicative of disease was identified (23).
Narrowband ultraviolet B radiation (nbUVB) is a common treatment for patients with moderate-to-severe psoriasis (24). UVB treatment of patients with atopic dermatitis has been shown to reduce skin-surface bacteria, mainly the S. aureus count, in several studies (25–27). UV radiation (UV-R) is known to trigger the production and release of antimicrobial peptides, affect the innate immune system, and ultimately suppress the adaptive cellular immune response (28, 29). However, so far, nothing is known about the effect of nbUVB treatment on the skin microbiome in patients with psoriasis.
The aim of this study is to analyse differences in the microbiome between lesional and non-lesional skin in patients with chronic plaque psoriasis and to investigate changes in the microbiome in these patients after nbUVB treatment. Furthermore, this study was designed to investigate to what extent responding to nbUVB treatment altered the skin microbiome.
A total of 30 patients with plaque type psoriasis with indication for nbUVB treatment were enrolled in this prospective study. Three patients did not complete nbUVB treatment and, for one patient, the microbiome samples failed, and were excluded from the study. Thus, 26 patients were included in the final data analysis (male:female ratio 20:6). None of the patients had received oral antibiotics, systemic anti-inflammatory or immune-modulating treatment 2 weeks prior to entering the study or while participating. Exclusion criteria included pregnancy, tanning or intensive sun exposure 2 weeks prior to enrolling in the study, being under the age of 18 years, known malignancy, psoriatic arthritis or other systemic inflammatory condition, as well as symptoms of active infection at the time of sample collection. Written informed consent was obtained from all subjects under protocols approved by the ethics committee Linköping University, Linköping, Sweden (approval number 2014/209-32). Sex, age, height, weight, current psoriasis treatment and current medications were recorded, as well as an assessment of disease severity using the Psoriasis Area and Severity Index (PASI), both from a designated target lesion (sampled lesional skin) and total body. Target lesion severity was calculated by adding the severity of erythema, induration and desquamation of the plaque (graded 0 = none to 4 = severe) and, since the area was excluded from the calculation, it was labelled target Psoriasis Severity Index (PSI). Participants were instructed not to apply topical anti-inflammatory treatment to the target lesion. For other lesions, topical treatment was restricted to moisturizing cream and mometasone 0.1%. The latter was applied to a maximum of 30% of the total body surface area. Patients’ demographic and clinical characteristics are shown in Table I. No control group of healthy individuals was analysed. This approach was chosen so as to not expose healthy people to UV-R, which increases the risk of skin cancer (30), and to avoid inter-individual variation in the skin microbiome (31).
Table I. Patients’ demographic data and Psoriasis and Area Severity Index (PASI) and target Psoriasis Severity Index (PSI) response after completed narrowband ultraviolet B (nbUVB) treatment
A sample from lesional skin (target lesion) and non-lesional skin at an adjacent cutaneous location at least 10 cm from the target lesion was taken from each patient by swabbing the skin in a 4×4 cm area with a flocked swab soaked in 1 ml liquid Amies (ESwab™, Copan Diagnostics Inc., Murrieta, CA, USA). Since the colonization of bacteria in the skin is dependent on the physiology of the skin site (32), all of the samples were taken from areas designated as dry microenvironments. The patients were treated with nbUVB according to clinic protocol and, after completing the treatment, new samples were taken from the same areas as before treatment and PASI and target lesion PSI was assessed again. Patients were advised to avoid additional sun exposure during daytime and not to visit tanning studios.
nbUVB (311nm) therapy was administered using a Waldmann 7002 cabin (Waldmann Medizintechnik, Villingen-Schwenningen, Germany). Patients were treated, on a mean ± standard deviation (SD) of 2.3 ± 0.7 times per week and the mean ± SD treatment period was 10.4 ± 3.6 weeks. The mean ± SD maximum dose reached was 2.64 ±1.2 J/cm2 at the end of the treatment period. Energy output was measured with a standard intrinsic UV meter. Initial dose was dependent on skin phototype. If the initial dose was tolerated, the previous dose was increased by 20% at each visit. When a previous treatment resulted in erythema, no treatment was given the next day, or the dose was decreased, depending on whether the erythema was asymptomatic or severe and painful.
The entire liquid sample suspension from the ESwab was transferred into 1.5 ml reaction tubes and centrifuged at 16,000 × g. The pellet was suspended in 180 µl G2 buffer (Qiagen, Hilden, Germany) and 20 µl Proteinase K (Qiagen) and incubated for 30 min at 56°C and then 5 min at 98°C. DNA extraction was performed in a Biorobot EZ1 (Qiagen) with the EZ1 Tissue kit v.2.0 (Qiagen) according to the manufacturer’s protocol. The sample volume for purification was 200 µl, and the purified DNA was eluted in 100 µl. A negative template control, one centrifuged E-swab, was prepared identically to the samples. Samples were prepared for 16S rDNA sequencing according to the 16S Metagenomic Sequencing Library Preparation guide part # 15044223 Rev. B. on a MiSeq instrument (Illumina, CA, USA). The protocol describes the steps to amplify the V3 and V4 region using a limited cycle PCR, to add Illumina sequencing adapters and dual-index barcodes to the amplicon target. Quality control was assessed using a DNA 1000 chip (Agilent Technologies, CA, USA) run in a Bioanalyzer 2100 (Agilent Technologies). DNA concentration was measured on a Qubit instrument with dsDNA HS Assay kit (Invitrogen, CA, USA). The only difference from the Illumina protocol was that the first PCR was duplicated and the same samples were then pooled before the first PCR clean up and that the negative template control was controlled after the first PCR on a Bioanalyzer DNA 1000 chip. All negative template controls were negative after the first PCR and therefore not further processed. Finally, 35% Phix Control and 6 pM amplicon library were used. Quality of sequencing was controlled by FastQC Java script (http://www.bioinformatics.babraham.ac.uk/projects/fastqc).
Fastq-files were imported as paired-end reads into CLC Genomic Workbench (http://www.clcbio.com version 9.5.1) for processing and analysis. Sequences were trimmed for quality (score 0.05, 0 ambiguities, discard <180 nt). Overlapping pairs were merged into a single sequence and merged sequences were further trimmed to remove PCR-primers, only keeping sequences ≥ 390 – ≤ 460 nt. Prior to clustering, all sequences were trimmed to a fixed length of 400 nt. The number of merged paired-end reads after quality filtering per sample varied from 6,792 to 330,650, with a mean number of reads of 76,950. Clustering was carried out using a reference-based approach, (Greengenes v.13.5 database (http://greengenes.secondgenome.com)) with a similarity threshold of 97%, and chimeric sequences were removed. The top 100 most abundant operational taxonomic units (OTUs) were aligned using MUSCLE with default settings and a maximum likelihood based phylogenetic tree was generated using neighbour joining and Jukes-Cantor nucleotide substitution model. Measures of alpha diversity (number of OTUs, Chao 1 bias-corrected, Shannon diversity index, Simpson’s index, and phylogenetic diversity) was calculated with a sample depth of 5,000, and visualized using alpha diversity rarefaction curves (20 points, no replacement, and 100 replicates at each point). Friedman analysis of variance (ANOVA) was used for statistical analysis of alpha diversity. Correlation between PASI/PSI and Shannon index was analysed using Spearman’s rank order correlation. Beta diversity was calculated using unweighted and weighted UniFrac, Bray-Curtis and Jaccard and a principal coordinates analysis (PCoA) was performed on the UniFrac distances. Significance testing was performed using the permutational multivariate analysis of variance (PERMANOVA) test in the CLC software.
Finally, the highly abundant taxa (mean abundance > 0.1% of total number of OTUs) were pairwise compared with regard to sample type and response to treatment at phylum and genus level using a generalized linear model, assuming read counts that follow a negative binomial distribution, and with a dispersion corrections similar to the EdgeR method (33). Significance testing was conducted using a false discovery rate (FDR) corrected p-value of 0.05.
Twenty-six participants received a mean of 21.2 UVB treatments (range 9–26) (Table I). The mean ± SD PASI before treatment was 8.0 ± 4.5 and after treatment 2.1 ± 1.8. Mean ± SD target PSI before treatment was 5.2 ± 1.4 and after treatment 1.1 ± 1.4. The mean ± SD PASI improvement was 69.9 ± 23.1% and target PSI improvement 76.5 ± 30.1%. Response to treatment was defined as achieving at least a 75% reduction in the PSI score of the target lesion from the start to the end of the trial, named target PSI75. Nineteen of the patients (73%) achieved target PSI75. Patients achieving target PSI75 were labelled responders and those not achieving target PSI75 non-responders.
Alpha diversity was measured using Shannon diversity index, Chao 1 estimator, Simpson’s index, and number of OTUs and no significant differences were found in the alpha diversity (Fig. 1A). PERMANOVA analysis of beta diversity (unweighted and weighted UniFrac, Bray-Curtis and Jaccard) revealed no significant difference between samples from lesional or non-lesional skin before and after treatment in responders or non-responders. Fig. 1B shows a PCoA of the beta diversity of lesional and non-lesional skin before and after nbUVB treatment. The samples do not cluster, indicating no similarity between the samples before or after nbUVB treatment.
Fig. 1. No significant differences in the alpha and beta diversity of microbiota before and after nbUVB treatment. (A) There is no significant difference in the alpha diversity in the skin microbiome in lesional and non-lesional skin before (pre) and after (post) narrowband ultraviolet B (nbUVB) treatment. Boxplot based on Shannon diversity index (n = 26, p > 0.05, • mean, box = mean ± 0.95 confidence interval, whiskers mean ± standard deviation). (B) Principal coordinate analysis (PCoA) of beta diversity. No significant clustering was seen in samples from lesional skin before (green) and after (blue) nbUVB treatment or in samples from non-lesional skin before (orange) and after (pink) nbUVB.
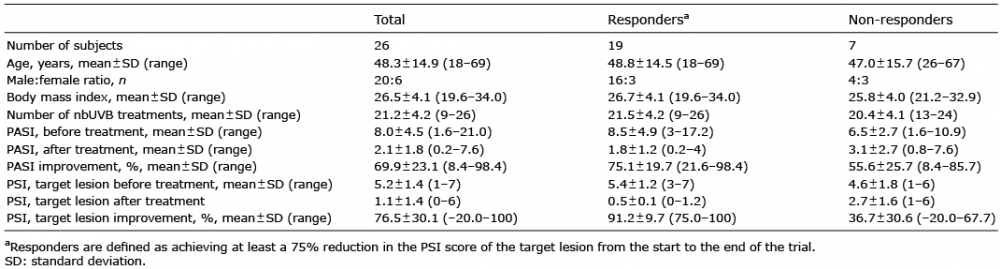
Our data showed a correlation between PASI before nbUVB treatment and Shannon diversity index (Fig. 2A, Spearman’s R = –0.55 and p < 0.01) and a correlation between target PSI and Shannon diversity index before treatment (Fig. 2B, Spearman’s R = –0.46 and p < 0.05). Our results revealed no correlation between PASI and Shannon diversity index after nbUVB treatment (R = –0.37 and p > 0.05) and no correlation between target PSI and Shannon diversity index after treatment (R = –0.09 and p > 0.05).
Fig. 2. Psoriasis severity is significantly correlated with alpha diversity. (A) Correlation between Psoriasis and Area Severity Index (PASI) before narrowband ultraviolet B (nbUVB) treatment (PASI pre) and Shannon index before treatment (Shannon index pre), (Spearman’s R=–0.55, p < 0.01). (B) Correlation between Psoriasis Severity Index (PSI) target lesion before nbUVB treatment and Shannon index pre (Spearman’s R=–0.46 and p <0.05) (n = 26, dotted line 0.95 confidence interval).
The majority of the bacteria fall into 3 dominating phyla: Firmicutes, Actinobacteria and Proteobacteria (Fig. 3A). Staphylococcus, Corynebacterium and Propionibacterium were the 3 most common genera (Fig. 4A). The significant differences found at the genus level when comparing lesional with non-lesional skin, pre-treatment with post-treatment, and responders with non-responders are summarized in Table II.
Fig. 3. Significantly decreased abundance of Firmicutes in lesional skin. (A) Bar chart of relative abundance of phylum level in lesional (L) and non-lesional (NL) skin before (pre) and after (post) narrow-band ultraviolet B (nbUVB) treatment. (B) Significant differences shown for Firmicutes (bar: mean + standard deviation, false discovery rate (FDR) p < 0.01=**, n = 26).
Fig. 4. Significant differences in the microbiome on genus level. (A) Bar chart of relative abundance of genus level in lesional (L) and non-lesional (NL) skin before (pre) and after (post) narrowband ultraviolet B (nbUVB) treatment (n = 26). (B) Bar chart of relative abundance of genus levels in lesional and non-lesional skin before (pre) and after (post) nbUVB treatment in target lesion responders (R) (n = 19) and non-responders (N) (n = 7). (C) Significant differences shown for Staphylococcus, Finegoldia, Anaerococcus, Peptoniphilus, Gardnerella, Prevotella and Clostridium (bar: mean + standard deviation, false discovery rate (FDR) p < 0.05=*. FDR p < 0.01=**).
Table II. Significant differences in relative abundance of genus levels in lesional and non-lesional skin before (pre) and after (post) narrowband ultraviolet B (nbUVB) treatment (n=26) and in target lesion responders (R) (n=19) and non-responders (N) (n=7)
Lesional skin had a significantly lower abundance of the phylum Firmicutes compared with non-lesional skin before UVB treatment (FDR p < 0.01, Fig. 4B) and no significant difference after treatment.
At the genus level, lesional skin had significantly lower levels of Staphylococcus (FDR p < 0.05), and higher levels of Conchiformibius (FDR p < 0.05) than non-lesional skin before nbUVB treatment. After treatment, lesional skin had significantly higher levels of Actinomyces (FDR p < 0.05) and lower levels of Paracoccus (FDR p < 0.05) compared with non-lesional skin.
nbUVB treatment led to an increase in the relative abundance of the genus Clostridium and a decrease in the relative abundance of the genus Pseudomonas in both lesional (FDR p < 0.01 and FDR p < 0.001) and non-lesional skin (FDR p < 0.05 and FDR p < 0.001), and an increase in the relative abundance of the genus Megasphaera in non-lesional skin (FDR p < 0.05).
Dividing patients into responders and non-responders revealed differences in the composition of the bacterial microbiome, shown in Fig. 5A for phylum level and Fig. 4B for genus level. Before nbUVB treatment, lesional skin had a significantly lower relative abundance of the phylum Firmicutes compared with non-lesional skin in non-responders, with no significant difference in responders (FDR p < 0.001, Fig. 5B). After nbUVB treatment, responders had significantly lower relative abundance of the phylum Firmicutes in both lesional (FDR p < 0.01) and non-lesional (FDR p < 0.001) skin (Fig. 5B) and significantly lower relative abundance of the phylum Bacteroidetes in non-lesional skin (FDR p < 0.05, Fig. 5C) compared with non-responders.
Fig. 5. Significant differences in the microbiome on phylum level in responders and non-responders. (A) Bar chart of relative abundance of phylum level in lesional (L) and non-lesional (NL) skin before (pre) and after (post) narrowband ultraviolet B (nbUVB) treatment in target lesion responders (R), achieving target PSI75, (R) (n = 19) and non-responders (NR), not achieving target PSI75, (n = 7). Significant differences shown for (B) Firmicutes, and (C) Bacteroidetes (bar: mean + standard deviation, false discovery rate (FDR) p < 0.05 = *. FDR p < 0.01 = ** and FDR p < 0.001=***). Cyanobacteria increased significantly in NL skin after treatment for the study group as a whole and in NR. The majority of the reads for Cyanobacteria were of the order Streptophyta, which is classified as a plant, not bacteria, and these results are therefore not regarded as relevant.
On genus level, before nbUVB treatment, responders had significantly lower relative abundance of Dermacoccus and Acinetobacter in both lesional (FDR p < 0.05 and FDR p < 0.01) and non-lesional skin (FDR p < 0.05 for both) and Staphylococcus in non-lesional skin (FDR p < 0.05). The nbUVB treatment led to a significant decrease in Pseudomonas in both lesional (FDR p < 0.001) and non-lesional skin (FDR p < 0.01) of responders with no significant difference in non-responders. After nbUVB treatment, responders had significantly lower levels of Staphylococcus, Finegoldia, Anaerococcus, Peptoniphilus, Gardnerella, Prevotella and Clostridium in lesional skin compared with non-responders (Fig. 4C).
Symbiotic interactions between the complex microflora and the skin have been shown to shape and modulate the innate immune response of the host (31). Maintenance of the right kind of microflora is thus believed to be essential for healthy skin. Several studies have connected modulation of cutaneous microbiome with allergic and inflammatory skin diseases, such as atopic dermatitis (19, 32). It is therefore tempting to speculate that altered interaction between microbiota and host or altered amounts of microbiota could be functional triggers for psoriasis.
The present study analyses, for the first time, differences in the microbiome between lesional and non-lesional skin of patients with psoriasis before and after nbUVB treatment. Our results showed 3 dominating phyla: Actinobacteria, Firmicutes and Proteobacteria, with Firmicutes being most prominent. This is in agreement with previous studies of the bacterial microbiome of normal skin. These phyla also constitute the main groups of microbiota found in the gastrointestinal tract and oral cavity, although in different proportions (32).
Gao et al. showed significantly higher relative abundance of Firmicutes in lesional compared with non-lesional skin (21), but, in our study, Firmicutes was significantly higher in non-lesional skin. It is difficult to compare the results, since Gao et al. did not describe the location of their skin sampling and they included fewer patients. Previous studies show that the colonization of bacteria in the skin is dependent on the physiology of the skin site, with specific bacteria being associated with moist, dry or sebaceous microenvironments. This makes it important that care is taken when comparing sites for microbiome studies (32). All the samples in the present study were taken from dry skin.
We found significantly lower levels of Staphylococcus in lesional skin compared with non-lesional skin before treatment, which is in contrast with the results of Tett et al.’s study (23) using shotgun metagenomics. They found that Staphylococcus was significantly more abundant on lesional skin, even when only comparing samples from the elbows, which is also regarded as dry skin. It is difficult to directly compare these studies from different geographical regions. Also, Tett et al. included patients receiving systemic treatments for their psoriasis and 4 of their patients had undergone a course of antibiotics in the last 3 months, which might have influenced results. Differences could also be due to what Yan et al. discuss in their review (34), that Staphylococcus is such a diverse genus that its role is probably better understood at species or even strain level. Some species, such as the commensal S. epidermidis, appears to enhance the innate immune barrier and limit pathogen invasion (35), while others, such as S. aureus, evoke a pathogenic Th17 response (36). Tett et al. found strains of S. epidermidis containing known virulence-related genes at psoriatic sites that were absent from strains at unaffected sites, and drew the conclusion that strain diversity is accompanied by potentially environmentally relevant functional differences that may be indicative of niche-specific selection with respect to psoriatic lesions (23).
Interestingly, the relative abundance of the genera Staphylococcus, Finegoldia, Anaerococcus, Peptoniphilus, Gardnerella, Prevotella and Clostridium, are all significantly lower in lesional skin of responders compared with non-responders after nbUVB treatment. Previous studies have shown that increased intestinal abundance of Prevotella copri correlates with an enhanced risk of developing rheumatoid arthritis (37), and that the skin commensal Finegoldia magna closely interacts with the skin immune system by inactivating antimicrobial peptides (38). There are also several previous studies suggesting an immune-modulating role of Clostridium. Levels of Clostridium leptum were decreased in IBD and spondyloarthritis (39, 40), and Kolho et al. (41) showed that children with IBD, responding well to treatment with tumour necrosis factor (TNF)-alpha inhibitors, developed increased relative abundance of Clostridium after treatment. In our study, Clostridium was not only significantly different between responders and non-responders. It also increased significantly, in both lesional and non-lesional skin, after nbUVB treatment, suggesting that immune-modulating effect of Clostridium is impacted by nbUVB treatment. We hypothesize that non-responders still have a high level of inflammation of the skin, in spite of treatment, which continually shifts the microbiome, leading to the changes between responders and non-responders we see after nbUVB treatment.
Other studies confirm that UV-R leads to various changes within the landscape of microbial communities of the skin. UV-R can reduce the growth of S. aureus in vivo and in vitro (26, 42, 43) and decrease the production of super-antigens, which are known to be potential triggers of immune responses (42, 44). UV-R is also known to induce production of antimicrobial peptides, which participate in innate immune responses and activate and mediate adaptive immune responses (28, 29).
In patients responding well to nbUVB, Pseudomonas decreased in both lesional and non-lesional skin. This is remarkable, since vitamin D3, which has a positive influence on gastrointestinal diseases, has been shown to decrease the relative abundance of Pseudomonas in the gut of healthy volunteers (45). The authors concluded that perhaps the treatment leads to a reduced inflammatory environment, which diminishes the competitive advantage of opportunistic pathogens, such as Pseudomonas, thus decreasing their abundance. This could also be true for our study. In non-responders, there is no significant change in Pseudomonas. Perhaps non-responders have a lower production of vitamin D in the skin in response to nbUVB and therefore there is still a high degree of inflammation after treatment. In the light of these results, it is tempting to speculate that the anti-inflammatory effect of topical vitamin D might decrease the abundance of Pseudomonas.
The relative abundance of Acinetobacter and Dermacoccus was significantly higher in both lesional and non-lesional skin of non-responders compared with responders before treatment. Acinetobacter has been found to be significantly more abundant in a Russian population compared with a Finnish population, leading the authors to the conclusion that this might contribute to the lower prevalence of allergies in Russia (46). In another study, Acinetobacter was far more abundant in lesional skin of patients with seborrheic dermatitis, compared with non-lesional skin (47). Dermacoccus has been found to be lower in atopic dermatitis-prone skin compared with normal skin (48). These studies suggest that Acinetobacter and Dermacoccus have a role in different inflammatory disorders. It can be speculated that a higher abundance of Acinetobacter and Dermacoccus is a negative prognostic factor for nbUVB treatment in psoriasis, but further studies are needed to confirm this.
The dysbiosis we see in our study in non-responders after nbUVB treatment might be a result of the lack of UVB efficacy or, more interestingly, the dysbiosis in itself could be responsible for the reduced effect of UVB. In that case, however, the dysbiosis would be expected to be present both before and after treatment, which our data does not support. We therefore hypothesize that the lack of response to UVB is responsible for the dysbiosis seen after treatment, which then occurs as a secondary phenomenon. Further studies, comparing other treatments for psoriasis, both topical and systemic, are needed to determine if they show the same pattern.
At the phylum level, there were also differences when comparing responders with non-responders. Firmicutes were significantly less abundant in lesional and non-lesional skin, and Bacteroidetes in non-lesional skin after treatment of responders compared with non-responders. It is difficult to interpret these changes in terms of clinical relevance, since each phyla consists of such a broad range of bacterial genera and species. Since previous studies have found the microbiome of psoriatic patients to be only discretely changed (49), we conclude that the changes at the genus level, and perhaps also at the species level, are more likely to be clinically significant.
The Shannon diversity indices of the samples in our study are congruent with indices found in another study (22). Alekseyenko et al. (18) found a trend towards decreased taxonomic diversity in psoriatic lesions, although non-significant, and Tett et al. found a shift towards decreasing richness in relation to disease status from samples from the ear, but not from the elbow skin area (23). Using the Shannon diversity index, we found no significant differences in alpha diversity between lesional and non-lesional skin. We did, however, find a correlation between severity of disease and alpha diversity before nbUVB treatment, where increasing PASI and target PSI relates to lower diversity. This correlation was not present after treatment. The severity of atopic eczema has been shown to correlate inversely with the diversity of the intestinal microbiome (50). Tett et al. found that species richness did not significantly correlate with disease severity (23) and Alekseyenko et al. studied whether specific taxa were correlated with PASI. They drew the conclusion that there is a weak link between clinical severity and microbial colonization (18).
Some of the patients achieved a 75% reduction in whole-skin PASI, but not in target lesion, and vice versa. We chose to relate our data to target PSI, since the treatment response and microbiome correlation in the target lesion is clinically more relevant than comparing with whole-skin PASI, and we chose achieving ≥ 75% improvement in PSI in the target lesion as the definition of treatment responders, since PASI75 is an established definition of treatment response. It would have been interesting to instead split into 3 groups: non-responders (target PSI < 50%), partial responders (target PSI 50–90%) full responders (target PSI > 90%), but more participants are needed for the groups to be large enough to draw further conclusions.
Limitations in the present study are the lack of a control group of healthy individuals. We decided not to include a control group, since the present study was conducted to analyse differences between lesional and non-lesional skin, and since nbUVB treatment increases the risk of skin cancer in healthy individuals. The structure of the cutaneous microbiome is dictated by the localized topography of the skin, and is generally observed to remain relatively stable over time (51–53). It has been shown that the skin microbiome differs more between subjects than in the same subject over time (51–53). It can, however, not be excluded that the time elapsed between samples or other unknown confounding factors contributed to the changes in the microbiome found in our study. Another study limitation is the male dominance in the included patients. Previous studies have shown inconsistent results when it comes to the contribution of sex to microbial communities (49). Some studies found differences in the relative abundances of numerous bacterial groups, and that women had higher levels of alpha diversity (54, 55), while other studies found no differences between bacterial communities in men and women (56, 57). It cannot be excluded that a more uniform sex ratio would have produced different results. Samples from matched sites on the same individual were analysed, making it less likely that the significant differences shown are a result of interpersonal variation.
In conclusion, the present study shows the first analysis of the effect of nbUVB treatment on the skin microbiome in psoriatic patients. We see several significant differences in the microbiome before and after nbUVB treatment, and between responders and non-responders to treatment. How-ever, further studies with more participants are needed to confirm our results. It would also be of great interest to study the effect of other anti-inflammatory treatments for psoriasis on the skin microbiome. If differences in the skin microbiota between responders and non-responders to specific treatment can be confirmed in larger studies, this could be used in the future to individualize treatment of psoriatic patients, leading to better treatment results.
This study was supported by grants from the Swedish Psoriasis Foundation and from Futurum, The Academy of Healthcare, County Council of Jönköping, Sweden. The authors are grateful for the help of Marita Skarstedt, Division of Medical Diagnostics and Siv Nilsson, Division of Dermatology, Region Jönköping County, Jönköping, Sweden.


 Click to show fullsize
Click to show fullsize Click to show fullsize
Click to show fullsize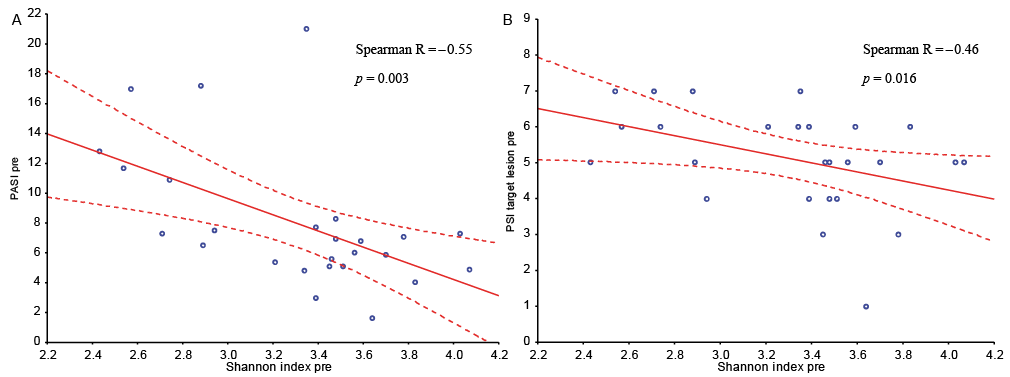 Click to show fullsize
Click to show fullsize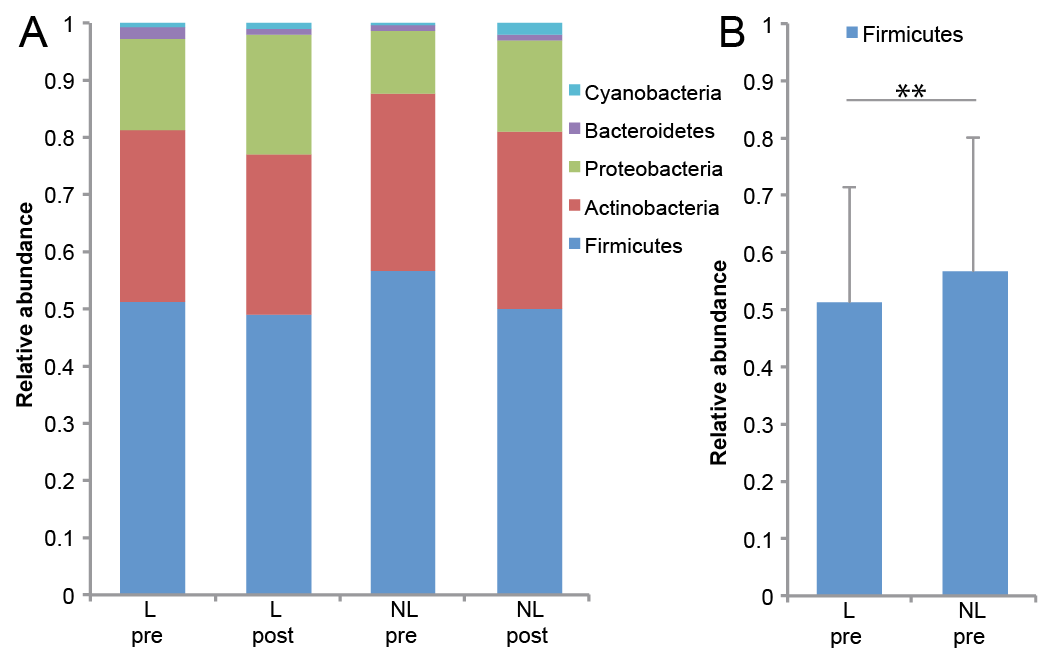 Click to show fullsize
Click to show fullsize Click to show fullsize
Click to show fullsize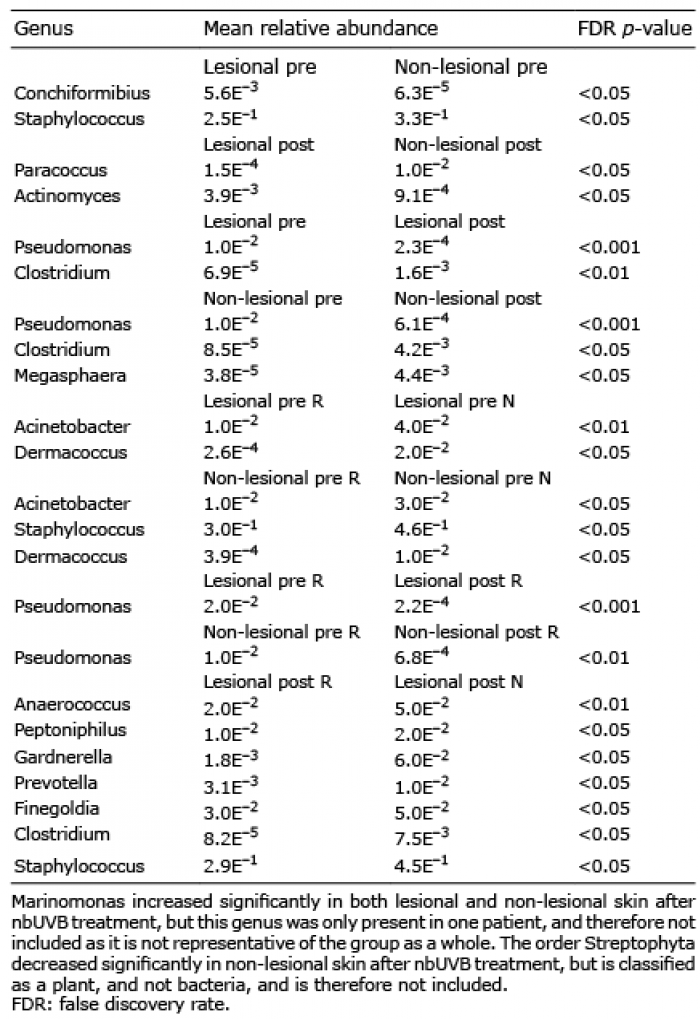 Click to show fullsize
Click to show fullsize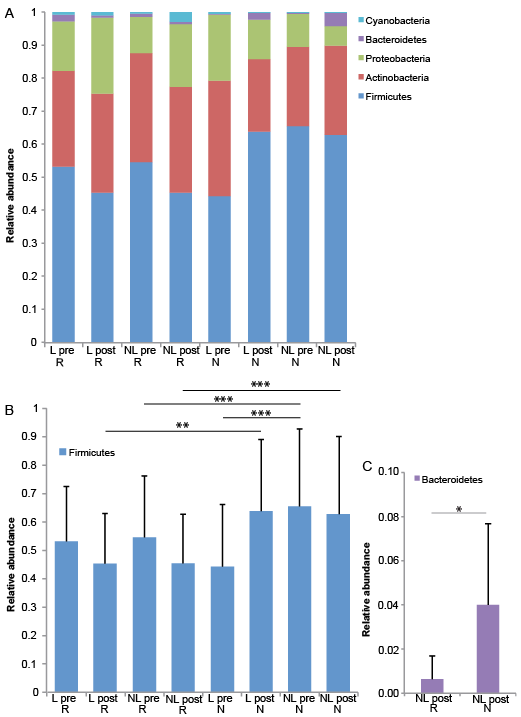 Click to show fullsize
Click to show fullsize