


Department of Dermatology, Affiliated Hospital of Guizhou Medical University, Guiyang, Guizhou, China
#These authors contributed equally to this paper.
Seborrhoeic keratosis is an age-related skin disease. Amyloid precursor protein (APP) plays an important role in the pathogenesis of age-related Alzheimer’s disease. The aim of this study was to elucidate the expression characteristics of APP in seborrhoeic keratosis tissues (n = 50), and explore whether the production of APP is related to the onset of seborrhoeic keratosis and skin ageing, including ultraviolet (UV)-induced ageing, as observed in normal skin (n = 79). The results of immunohistochemistry, Western blotting and quantitative real-time PCR showed that APP and its downstream products (i.e. amyloid-β42) were more highly expressed in seborrhoeic keratosis than in paired adjacent normal skin tissues. In contrast, the expression of its key secretase (i.e. β-secretase1) was generally low. Furthermore, APP expression was higher in UV-exposed than non-exposed skin sites, and expression in the older age group (61–85 years) was greater than that in the younger age group (41–60 years) in seborrhoeic keratosis tissues (p<0.05). APP expression correlated positively with age in epidermis (p<0.05), but not in dermis. These findings suggest that overexpression of APP may promote the onset of seborrhoeic keratosis and is a marker of skin ageing and UV damage. Further research will elucidate whether therapeutic mitigation of increased levels of APP in the skin might delay the onset of seborrhoeic keratosis and skin ageing.
Key words: amyloid precursor protein; seborrhoeic keratosis; amyloid-β42; β-secretase1 (BACE1).
Accepted Feb 20, 2018; Epub ahead of print Feb 28, 2018
Acta Derm Venereol 2018; XX: XX–XX.
Corr: Hongguang Lu, Department of Dermatology, Affiliated Hospital of Guizhou Medical University, Guiyang, Guizhou 550001, China. E-mail: hongguanglu@hotmail.com
Seborrhoeic keratosis (SK) is one of the most common benign epithelial skin tumours. SK is caused by the abnormal proliferation of epidermal keratinocytes (KCs) (1). SK lesions are often round or oval-shaped, yellowish, grey-brown or black, usually elevated, and appear to be stuck on the skin surface. They may arise on any area of the skin surface, except for the palms and soles (1, 2). Histologically, SK is composed mainly of basaloid and squamous cells (3). Both UV exposure and old age are independent risk factors for SK (1), which is often considered as a sign of skin ageing, including photoaging (4). SK is causally related to somatic mutations in FGFR3 and a few other genes expressed in human keratinocytes (5). Like senile Alzheimer’s disease (AD), the prevalence of SK tends to increase with age (1).
It is well-known that amyloid precursor protein (APP) is a key player in the pathogenesis of senile AD. APP is an integral type I transmembrane protein, which contains at least 19 exons that are located on chromosome 21 (21q21.2-3) (6, 7). The cDNA sequence of human APP was first cloned from a brain tissue library (6, 8). APP is processed by at least 3 proteases, i.e. α-, β- and γ-secretases, via the amyloidogenic and non-amyloidogenic pathways (9). In the amyloidogenic path-way cleavage by β-secretase (BACE) and γ-secretase releases amyloid-β42 (Aβ42), which plays an important role in early pathological events in early-onset familial AD and sporadic age-related AD (8, 10). Moreover, previous studies have shown that other age-related diseases, such as Parkinson’s disease and age-related macular degeneration, are accompanied by high levels of expression of APP (11–14). However, the biological activity of APP in SK is unknown. There is substantial evidence to support that APP is expressed not only in neuronal cells of the brain, but also in non-neuronal cells, such as KCs (7, 15–18). In skin tissue, researchers have observed that APP is highly expressed within the basal cell layer of mammalian epidermis (9). Functionally, both full-length APP and the soluble form (sAPPα), cleaved by α-secretase via the non-amyloidogenic pathway, can enhance proliferation, differentiation and migration of KCs (9). Although APP is up-regulated in some proliferative skin diseases, such as psoriasis (9), it remains unclear whether expression of APP is associated with the development of age-related skin diseases and skin ageing.
The aim of this study was to examine the expression of APP, BACE1 and Aβ42 in SK, and to elucidate the relationship between expression of APP and skin ageing.
A total of 129 subjects (50 patients with SK and 79 healthy controls) were recruited at the Department of Dermatology, Affiliated Hospital of Guizhou Medical University between January 2015 and July 2016. SK tissues and paired adjacent normal skin tissues (PANST) were collected from the 50 patients with SK. Diagnoses of all patients were based on clinical examination and skin biopsy by 2 experienced dermatologists according to diagnostic criteria for SK (1). Normal skin tissues from different parts of the body were obtained from 79 subjects without SK, who were undergoing plastic surgery. The characteristics of all samples are shown in Table I. The areas of UV exposure and non-exposure were outlined according to a previous publication (19). This study was approved by the ethics committee of the medical faculty in Guiyang. All subjects provided written informed consent.
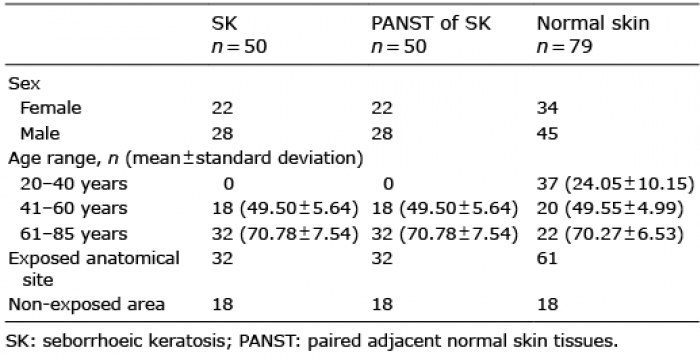
Table I. Participants’ characteristics
The samples of SK tissues included SK lesions and 0.5–0.7-cm PANST. The fresh samples were rinsed 3 times with sterile phosphate buffered saline (PBS) and the subcutaneous tissues were excised from above the samples using ophthalmic scissors. Adjacent normal skin samples were then separated from SK lesions for further study. The normal skin tissues were divided into 2 parts, and the epidermis and dermis were separated from one part. Each of the normal skin tissues was cut into small tissues (2×2 cm) and the epidermis was separated from the dermis after incubation with 0.25% dispase for 16–18 h at 4°C.
All of the above samples were prepared for Western blotting (WB), quantitative real-time PCR (qRT-PCR) and immunohistochemistry staining.
Fifty biopsies including SK tissues and PANST were available. The skin tissue samples were fixed in 10% formalin solution at 4°C for 24 h. Paraffin embedding, sectioning (4-μm thick sections) and routine haematoxylin and eosin staining were performed at the Department of Pathology of Affiliated Hospital of Guizhou Medical University. The diagnoses were confirmed with light microscopy (Leica DM2000, Wetzlar, Hessen, Germany) by 2 dermatopathologists. Immunohistochemistry (IHC) of 23 SK and PANST samples was performed according to a previous reference (20). Mouse monoclonal anti-APP (1:150, Abcam, Cambridge, UK), rabbit polyclonal anti-Aβ42 (1:150, Abcam) and mouse monoclonal anti-BACE1 (1:150, Santa Cruz Biotechnology, Santa Cruz, CA, USA) antibodies were used as primary antibodies. Those paraffin sections were incubated for 30 min with the above primary antibodies. The samples were then rinsed twice with PBS, incubated with horseradish peroxidase-conjugated IgG goat anti-mouse or rabbit secondary antibody (1:150, DAKO, Glostrup, Denmark) for 30 min and visualized with 3-amino-9-ethylcarbazole (DAKO). Negative controls were prepared by replacing the primary antibody with PBS. The sections were counterstained with haematoxylin and observed under the light microscope (Leica DM2000, Germany).
For WB analysis, SK tissues, PANST and normal skin tissues (including the epidermis, dermis and the entire skin tissues) from different age groups were used as described previously (21). Total protein extracts were obtained by tissue lysis in RIPA lysis buffer (Solarbio, Beijing, China) containing 1 μm PMSF (Beyotime Biotechnology, Shanghai, China). For WB, 40 μg protein was subjected to SDS-PAGE and then transferred onto nitrocellulose membranes (Beyotime Biotechnology, Shanghai, China). Membranes were blocked with non-fat milk for 2 h at room temperature and then incubated with primary antibodies overnight at 4°C. In the WB assay, the above mouse monoclonal anti-APP antibody (1:1000, Proteintech Group, Wuhan, Hubei, China), rabbit polyclonal anti-Aβ42 antibody (1:1000, Abcam), mouse monoclonal anti-BACE1 antibody (1:1000, Santa Cruz Biotechnology, Santa Cruz, CA, USA) were used as primary antibodies. Anti-β-actin antibody (1:2000, Beyotime Biotechnology) was used as an internal control. After washing 4 times with Tris-buffered saline Tween washing buffer (TBST buffer), the blots were incubated with horseradish peroxidase-conjugated anti-mouse or anti-rabbit IgG (1:1000, DAKO) for 2 h at room temperature. Finally, expression of the above proteins was detected using ECL WB detection reagent (7sea Biotechnology, Shanghai, China) after washing 6 times with TBST buffer. The detection of protein expression was repeated 3 times, and its expression levels measured with FastStone Image Viewer 5.5 software after normalization with β-actin.
qRT-PCR was performed as described previously (22). Total RNA was isolated from SK tissues, PANST of SK, and normal skin tissues (including the epidermis, dermis and the entire skin tissues) from different age groups by Trizol reagent (Invitrogen, Paisley, Renfrewshire, UK). The concentration and quality of extracted RNA in each sample was tested using the Epoch Microplate Spectrophotometer (BioTek Instruments Inc., Winooski, USA). RevertAid first-strand cDNA synthesis kit (Invitrogen) was used to synthesize first-strand cDNA from 0.3 μg total RNA with random hexamer primers according to the manufacturer’s instruction. qRT-PCR analysis was performed using an Eppendorf system (Realplex, Hamburg, Germany) with SYBR Green Supermix (Tiangen, Beijing, China) in the amplification reaction mixtures (25 μl). The primer sequences were: APP: 5’-CTACCGCTGCTTAGTTGGTGAGT-3’ (forward), 5’-TGTGCCAGTGAAGATGAGTTTCG-3’ (reverse); Aβ42: 5’-AACAGAAGGACAGACAGCA-3’ (forward), 5’-TCATAAATCACACGGAGGT-3’ (reverse); BACE1: 5’-GGCGGGAGTGGTATTATGAG-3’ (forward), 5’-GGCTGCCTTGATGGATTTGA-3’ (reverse); GAPDH: 5’-GAGAAGTATGACAACAGCCTCAAGATCA-3’ (forward), 5’-CATGAGTCCTTCCACGATACCAAA-3’ (reverse).
The reaction was carried out by pre-degeneration at 95°C for 10 min, 40 cycles at 95°C for 10 s, 55°C for 30 s and 72°C for 32 s. All samples were tested in triplicate, with the reference gene GAPDH for data normalization to adjust the variations in RNA quantity and quality. Relative mRNA level was calculated using the 2-?CT method.
Data were analysed using GraphPad Prism 5 statistical software using the Wilcoxon rank sum test. A p-value < 0.05 was considered significant.
IHC analyses of 23 samples of SK and PANST, respectively, showed that APP was expressed in the entire epithelial layer in SK tissues (Fig. 1a), but only moderately expressed in the basal cell layer in the adjacent skin (PANST) (Fig. 1b), i.e. similar to previous findings in normal skin (8). Consistent with these results, WB analysis showed that APP was highly expressed in SK tissues compared with PANST (Fig. 2a).
Fig. 1. Expression of amyloid precursor protein (APP) in seborrhoeic keratosis (SK) tissues and paired adjacent normal skin tissues (PANST) analysed by immunohistochemistry. (a) APP emitted strong perinuclear staining in the entire SK tissue, which was probed with anti-APP antibody. (b) APP was mainly expressed in the epithelial basal cell layer in PANST (a, b: left ×100, right ×400).
Fig. 2. Expression levels of amyloid precursor protein in seborrhoeic keratosis (SK) tissues (n = 50) and paired adjacent normal skin tissues (PANST) (n = 50) detected by Western blotting (WB). (a) APP expression in SK tissues (SK1, SK2) was significantly higher than PANST (PANST1, PANST2) (left panel). APP was analysed by WB with anti-APP antibody and β-actin. Results were quantified by comparing the densitometric values (right panel). (b) APP expression in the exposure area (n = 32) of SK tissues was higher than in the non-exposed area (n = 18). (c) APP expression in the 61–85-year group (n = 32) was significantly higher than in the 41–60-year group (n = 18). Data represent mean ± SD of APP level normalized to that of β-actin and relative to that of PANST; *p < 0.05.
To explore the role of UV exposure and skin ageing on expression of APP, we compared SK tissues from UV-exposed and non-exposed skin sites from 2 age groups. APP was highly expressed in UV exposure sites compared with non-exposure sites (Fig. 2b), and the expression level was higher in the 61–85-year group than in the 41–60-year group (p < 0.05) (Fig. 2c). However, there was no significant difference between women and men (data not shown).
The expression levels of APP mRNA analysed by qRT-PCR were consistently higher in SK tissues compared with PANST (Fig. 3a). Moreover, the expression levels in SK tissues from UV exposure sites were higher than in non-exposure sites (Fig. 3b), and the levels in the older patients were higher than those in the younger patients (Fig. 3c).
Fig. 3. Expression levels of amyloid precursor protein (APP) mRNA in seborrhoeic keratosis (SK) tissues (n = 50) and paired adjacent normal skin tissues (PANST) (n = 50) detected by quantitative real time-PCR. (a) Expression level of APP mRNA in SK tissues was significantly higher than PANST. (b) Expression level of APP mRNA in the exposure area (n = 32) of SK tissues was higher than in the non-exposed area (n = 18). (c) Expression of APP mRNA in the 61–85-year group (n = 32) was significantly higher than in the 41–60-year group (n = 18). Data represent mean ± SD of APP levels normalized to that of GAPDH. *p < 0.01.
To clarify whether APP expression was associated with skin ageing, we detected the protein and mRNA expression of APP in tissue samples from different age groups of volunteers without SK. Total protein and mRNA were extracted from non-separated normal skin tissues and separated epidermis and dermis from different age groups. APP and mRNA expressions were examined by WB and qRT-PCR, respectively. The expression levels of APP and mRNA were positively correlated with ageing in epidermis (Fig. 4a, b), but not in dermis and non-separated normal skin (data not shown).
Fig. 4. Expression of amyloid precursor protein (APP) and mRNA levels increased with age in normal epidermis, as detected by Western blot (WB) and quantitative real time-PCR. (a) Expression level of APP increased significantly with age, as evidenced by WB using anti-APP antibody and β-actin. Results were quantified by comparing the densitometric values. Data represent mean ± SD of APP level normalized to that of β-actin. p < 0.001. (b) The expression level of APP mRNA in the epidermis of normal skin tissues increased with age. Data represent means ± SD of APP levels normalized to that of GAPDH. p < 0.01. 20–40-year age group (n = 37); 41–60-year age group (n = 20); 61–85 year age group (n = 22).
We further examined the protein and mRNA expression levels of Aβ42 and BACE1in SK tissues by IHC, WB and qRT-PCR, and the results were consistent. The protein and mRNA expressions of Aβ42 in SK tissues were higher than those in PANST (Fig. 5a, b and Fig. 6a, c). Less BACE1 protein and mRNA were expressed in SK tissues than in PANST (Fig. 5c, d and Fig. 6b, d).
Fig. 5. Immunohistochemistry characteristics of amyloid-β42 (Aβ42) and β-secretase1 (BACE1) in seborrheic keratosis (SK) tissues and paired adjacent normal skin tissues (PANST). (a, b) The protein expression of Aβ42 in the entire SK tissue was higher than that in PANST. (c, d) The protein expression of BACE1 in SK tissues was lower than that in PANST (a–d: left ×100; right ×400).
Fig. 6. (a, b): Protein expression of amyloid-β42 (Aβ42) and β-secretase1 (BACE1) in seborrhoeic keratosis (SK) tissues (n = 50) and paired adjacent normal skin tissues (PANST) (n = 50) examined by Western blot (WB). (c, d): mRNA expression levels of Aβ42 and BACE1 were determined in SK tissues (n = 50) and PANST (n = 50) by quantitative real time-PCR. (a, c): Aβ42 protein and mRNA were highly expressed in SK tissues compared with PANST. (b, d): BACE1 protein and mRNA were lowly expressed in SK tissues compared with PANST. p < 0.05.
As an age-related disease, SK is one of the manifestations of skin ageing (23). Since it is a benign epithelial tumour treatment is not mandatory, although surgical removal is not uncommon, especially in those with multiple lesions, both for cosmetic reasons and when the clinical diagnosis is uncertain. In previous studies APP was found to be associated with some age-related nervous system diseases (8, 10–14), but this does not appear to have been investigated in the context of skin disorders and skin ageing. In the skin, APP is mainly expressed in keratinocytes and melanocytes of the epidermal basal layer, with a small amount in dermal fibroblasts of the epidermal–dermal junction (9, 24, 25). We herein found that APP was highly expressed in the whole epithelial SK tissue.
Structurally, APP contains a growth factor-like globular domain rich in cysteine (26). As a surface receptor in epidermal KCs, it predominantly participates in protecting the cells, inhibiting their apoptosis, and enhancing their proliferation and migration (9, 18, 24, 25, 27). Moreover, similar to growth factor and tumour growth factor alpha (TGF-α), APP can induce proliferation of epithelial KCs (18, 28). Furthermore, a critical role of epidermal growth factor receptor (EGFR) has been identified in skin ageing, and EGFR is up-regulated in SK (29, 30). EGFR is essential to the biology of epidermal KCs, and APP also participates in regulating EGFR (7). EGFR mutations and up-regulations have been detected in different kinds of tumour, including SK (7, 31). Interestingly, APP can mediate melanosome transport, and sAPPα can promote the exocytic release of melanin by melanocytes via the autocrine mode or paracrine mechanism from neighbouring KCs (9), indicating a role of APP in the pathogenesis of pigmented SK lesions.
In addition, we herein demonstrate higher expression of APP in UV-exposed areas of SKs than in the non-exposure area, and that such expression is higher in the 61–85-year-old group than in the 41–60-year-old group. We further found that the expression of APP in the epidermis of normal skin tissues increased significantly with age, but that such expression in the dermis tissues did not (data not shown). APP has been reported to be a marker for cellular senescence (32–34), and it is highly expressed or mutated in some age-related diseases, such as AD, atherosclerosis, Parkinson’s disease and age-related macular degeneration (8, 11–14). Skin ageing is accompanied by many functional decrements, including mitochondrial energy metabolism and responsiveness of cells mitogens, being related to age-associated changes and damage due to environmental factors, such as UV radiation (35). Mitochondrial damage accelerates ageing through mitochondrial dysfunction, mtDNA and accumulation of oxidative damage over time as a result of elevated reactive oxygen species (36, 37). Up-regulation of APP contributes to mitochondrial dysfunction and decreases the expression of the main metabolic enzymes in the tricarboxylic acid cycle of mitochondrial and oxidative phosphorylations (38–40). In addition, SK risk factors, including oxidative stress, abnormal lipid metabolism, and abnormal glucose metabolism, will increase with organism ageing, which are also associated with APP (41). APP increases with age in a senescence-accelerated mouse model and Ts65DN Down syndrome mouse model (33, 34). Therefore, the activation of APP is correlated with UV exposure and age. Taken together, APP is an age-dependent protein and can be considered as a marker of skin ageing.
Much evidence implicates generation of amyloid-β from cleaved APP as a critical component of AD pathogenesis, especially for Aβ42 (8, 10). We sought to investigate the characteristic of Aβ in SK tissue. The production of Aβ is much more complicated than expected, and probably occurs via multiple pathways (42). As a main APP cleavage enzyme, BACE1 is rate-limiting for the generation of Aβ (10, 43). In our study, Aβ42 was overexpressed in SK tissues and BACE1 was low. The high expression of Aβ42 may be attributed to BACE1 competitive secretory enzymes which cleaved APP. However, the pathway related to Aβ42 release has not been entirely elucidated by cleaving APP. Liu et al. reported that Aβ42 mRNA expression in choroid plexus epithelial cells increased with age, and Aβ accumulated because the expression of Aβ transporter-associated protein reduced, leading to oxidative stress injury (44). Amyloid was also found to be deposited in SK tissues, but it is not clear whether it is Aβ42 (45, 46). We herein found that APP expression in SK tissues may be regulated by BACE1 and Aβ42, and may contribute to the onset of SK.
In summary, this study demonstrates, for the first time, that APP is highly expressed in SK tissues. APP expression in the epidermis of normal skin tissues increased with age. APP may be a biomarker of skin ageing, but whether this is a primary event, or is secondary to other processes involved in skin ageing and SK pathogenesis, needs further research.
The authors would like to thank Professor Jishi Wang’s (Key Laboratory of Hematological Disease Diagnostic & Treat Centre of Guizhou Province) research team for skilful technical support, and Dr Anran Fan (Tissue Engineering and Stem Cell Experiment Center, Guizhou Medical University) for language editing.
Funding support: National Natural Science Foundation of China (No. 81673069).
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize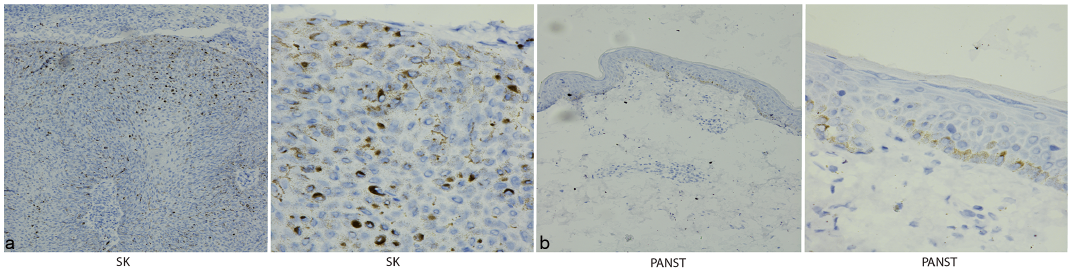 Click to show fullsize
Click to show fullsize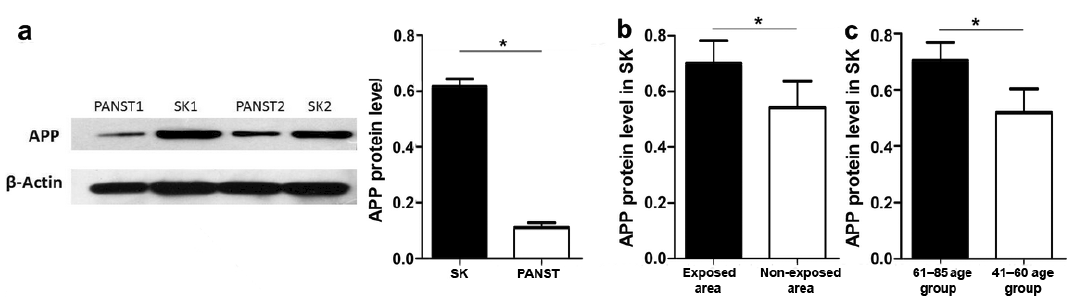 Click to show fullsize
Click to show fullsize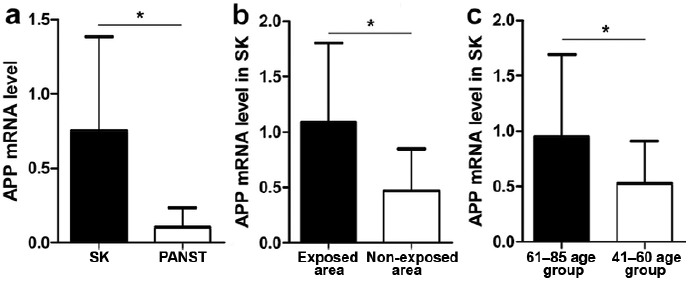 Click to show fullsize
Click to show fullsize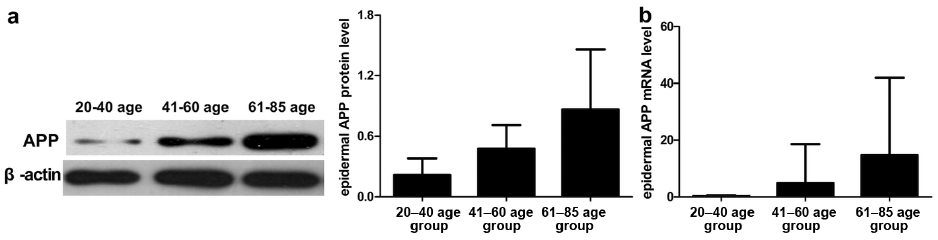 Click to show fullsize
Click to show fullsize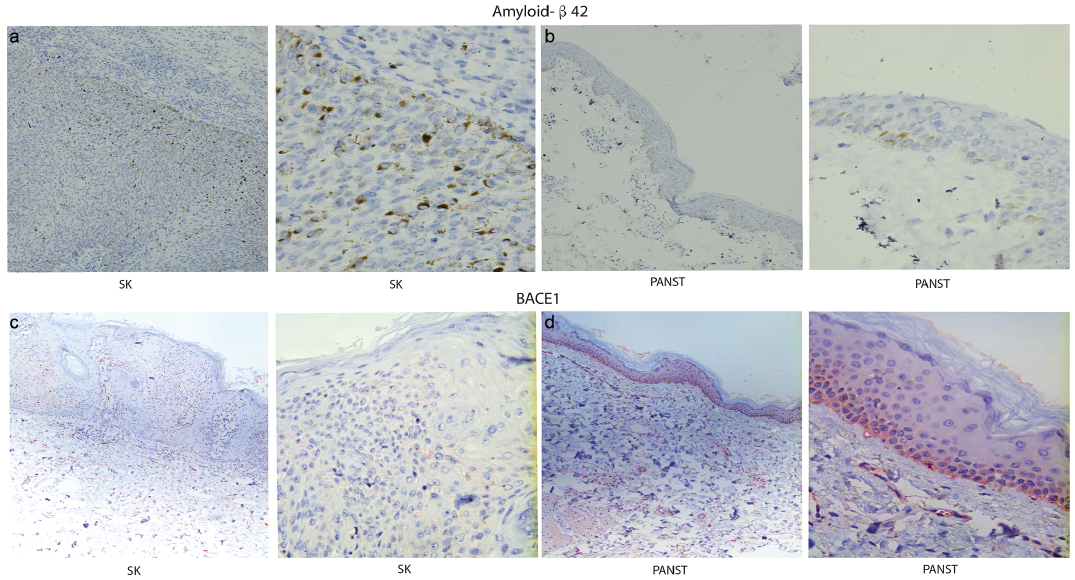 Click to show fullsize
Click to show fullsize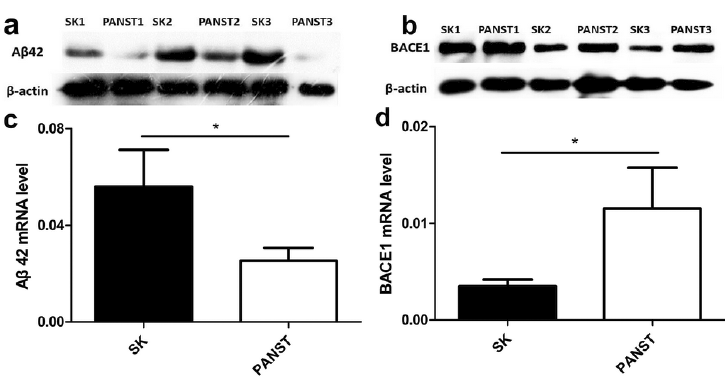 Click to show fullsize
Click to show fullsize