


1Department of Virology and 4Spatial Foodweb Ecology Group, The Faculty of Agriculture and Forestry, University of Helsinki, PO Box 21, FIN-00014 Helsinki, 2Department of Dermatology, Allergology and Venereal Diseases, University of Helsinki and Center of Inflammation, Helsinki University Hospital, Helsinki, and 3Biodiversity Unit, University of Turku, Turku, Finland. E-mail: Kati.Hokynar@helsinki.fi
Accepted Jun 26, 2018; Epub ahead of print Jun 29, 2018
Chlamydia-related bacteria represent a diverse group of bacteria within the order Chlamydiales that have been detected ubiquitously in various environmental samples, such as soil and water, in animals, and also in humans (1–6). We recently demonstrated the presence of Chlamydiales DNA in human skin (7). Chlamydiales DNA could be detected in > 50% of all the skin biopsies studied, but the prevalence was higher and the type range was wider in biopsies from individuals with a suspected tick-related skin condition, compared with biopsies from healthy skin (8).
Parapsoriasis is part of the continuum of cutaneous lymphoproliferative disorders of unknown aetiology and is clinically classified into small-plaque parapsoriasis (SPP) and large-plaque parapsoriasis (LPP) (9). This study examined the prevalence, distribution and relative abundance of Chlamydiales DNA in skin biopsies and skin swab samples, both from lesional and contralateral healthy skin, from 13 patients with parapsoriasis (6 SPP, 7 LPP; see Appendix S1). The clinical characteristics and skin microbiome data for these patients have been reported earlier (10).
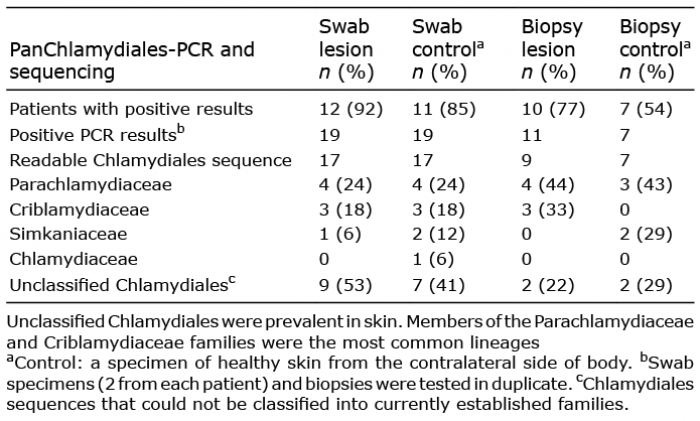
No Chlamydiales sequences were detected among the previous microbiome data (10) when searched specifically for Chlamydiales sequences (Appendix S1). Nonetheless, using a Chlamydiales-specific method, 12/13 (92%) patients with SPP or LPP had Chlamydiales DNA in the skin swabs taken from the lesional site and 11 (85%) in swabs from the contralateral healthy skin. The prevalence of Chlamydiales DNA was lower in the biopsy specimens than in the swab specimens: 77% and 54% of the patients had Chlamydiales DNA in the biopsies of lesional and control skin areas, respectively.
In swabs from both lesional and healthy skin, the majority of the Chlamydiales sequences remained unclassified (Table I and Table SI). In BLAST analysis, Parachlamydiaceae (24%), Criblamydiaceae (18%) and Simkaniaceae (6–12%) were the closest established Chlamydiales families (with ≥ 90% identity) for sequences present in swabs from both healthy skin and parapsoriasis lesions (Table I and Table SI). In skin biopsies, Parachlamydiaceae sequences were detected in both healthy and parapsoriatic skin (up to 44% of all sequences), while Criblamydiaceae sequences were detected only in lesional (33%) and Simkaniaceae (29%) only in healthy skin biopsies. No difference in the sequence heterogeneity was observed between SPP and LPP. Interestingly, using sequence analysis, heterogeneous Chlamydiales sequences were seen in the duplicate samples (both swabs and biopsies), suggesting that several types of Chlamydiales can be present simultaneously in skin. The ΔCt values (Ctpanchlamydiales – Ctbetaglobin) were smaller in swabs than in biopsies, indicating that the relative quantity of Chlamydiales DNA was higher in swab samples than in biopsy samples (p < 0.0001) (Fig. S1).
Table I. Chlamydiales families in the sequenced pan-Chlamydiales PCR-positive skin samples
This study confirmed that skin contains detectable amounts of Chlamydiales DNA. Earlier, we showed that skin microbiomes in parapsoriasis lesions and healthy skin were very similar, suggesting that parapsoriasis is not associated with alterations in the composition of patient’s cutaneous bacterial community, but interpersonal variation was significant (10). In that study, Chlamydiales DNA was not detected, as the 16S rRNA gene sequences of the Chlamydiales order differ from those of other bacteria, and primers used in pan-bacterial PCRs often fail in detection of Chlamydiales (2). Indeed, Chlamydiales DNA was frequently detected in skin samples, when Chlamydiales-specific primers were used. It is also possible that the quantity of Chlamydiales sequences is low and outnumbered by the other bacteria.
In the swab specimens, both the prevalence and relative quantity of Chlamydiales DNA were higher than in skin biopsies. No significant differences in the occurrence of Chlamydiales DNA or in the sequence heterogeneity were observed between the swabs taken from skin lesions and those from healthy skin, whereas in skin biopsies from the lesions (77%) Chlamydiales were more often detected than in biopsies of healthy skin (54%). Similarly, in our earlier studies, the prevalence of Chlamydiales DNA was >70% in biopsies from various skin lesions (tick-bitten skin, granuloma annulare and morphea), whereas approximately half of the biopsies of healthy skin harboured Chlamydiales sequences (7, 8). The slightly higher prevalence seen in the current study might be due to differences in storage conditions of the samples.
This unique sample material gave us an opportunity to study whether certain types of Chlamydiales could be found in healthy skin, or concentrated in the inflammatory lesions (parapsoriasis lesional skin), and whether some Chlamydiales could be detected on the skin surface (swabs) and others in the deeper layers of skin (biopsies). The diversity of Chlamydiales sequences in skin swabs and biopsies of parapsoriasis resembled that of healthy skin and morphea described in our earlier studies (7, 8): in all these groups, the majority of classified sequences belonged to Parachlamydiaceae, Criblamydiaceae and Simkaniaceae families. Consequently, these Chlamydiales are not likely to play a significant role in the pathogenesis of parapsoriasis. Neither the human pathogen Chlamydia trachomatis nor Chlamydia pneumoniae were present in skin. However, there were a relatively large proportion of Chlamydiales sequences that could not be classified with the current classification scheme (7, 11, 12), suggesting that as-yet unknown chlamydial families might be present in skin.
The origin of Chlamydiales in skin specimens remains obscure. In our earlier studies, Rhabdochlamydiaceae and Waddliaceae sequences were detected in ticks and in tick-related skin manifestations (7). In skin swabs and biopsies of the patients with parapsoriasis, the Chlamydiales families detected resembled those observed in normal skin, and members of the Rhabdochlamydiaceae and Waddliaceae were not found. In addition to ticks, free-living amoebae and various water sources, such as domestic drinking/shower water can harbour Chlamydiales bacteria (13–15). Members of Paracahlamydiaceae, Criblamydiaceae, Waddliaceae and Simkaniaceae have been identified in water, and thus it can be speculated that Chlamydiales DNA detected on the skin surface might originate from water sources (13, 14).
In conclusion, Chlamydiales DNA is frequently detected in skin samples, but is not likely to play a direct role in the pathogenesis of parapsoriasis. Nevertheless, the transmission routes and pathogenic potential of Chlamydiales in skin diseases warrant further study.
The authors are grateful to Dr Petri Auvinen for providing extracted DNA from the skin swab samples. The authors also thank Alli Tallqvist and Anu Kaitonen for technical assistance. This study was supported by the Academy of Finland (project #1285975), Jane and Aatos Erkko Foundation, and by research grants from Helsinki-Uusimaa Hospital District (TYH2015318/MP TYH2017244/MP). The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize