


1Department of Dermatology, Daejeon St Mary’s Hospital, The Catholic University of Korea, Seoul, Korea, and 2Department of Dermatology, The Saarland University Hospital, DE-66421 Homburg/Saar, Germany. E-mail: joerg.reichrath@uks.eu
Accepted Feb 10, 2020; Epub ahead of print Feb 13, 2020
Acta Derm Venereol 2020; 100: adv00098
Basal cell carcinoma (BCC) is the most common malignant tumour of the skin. The incidence of BCC is increasing, and the disease represents a major medical and economic problem. UV radiation and other genotoxic agents have been shown to cause different types of both direct and indirect DNA damage (1), including the generation of pyrimidine dimers and DNA double-strand breaks (DSBs). In addition to mutations in the sonic hedgehog pathway, which represent the molecular hallmark of BCC pathogenesis, mutations in the p53 gene are frequently detected in these malignancies. However, the relevance of this finding is not completely understood; thus, the aim of this investigation was to elucidate the relevance of the p53 pathway in the pathogenesis of BCC by immunohistochemically analysing the expression of some major proteins involved in p53-mediated cellular signalling, namely p53, ?Np63, TAp73, and MDM2. Because p53 is activated by DNA damage to either arrest the cell cycle or induce apoptosis, we also immunohistochemically analysed the expression of γ-H2AX, an early sensor of DNA damage.
Biopsy specimens that had been diagnosed as BCC by a certified pathologist were collected from the Clinic for Dermatology, Venereology and Allergology of the Saarland University Hospital in Homburg, Germany. Paraffin sections (5 μm) of BCC specimens (n = 10) were immunohistochemically analysed by applying a streptavidin-peroxidase technique using DAKO REAL detection system (alkaline phosphatase/RED rabbit/mouse, cat. no. K5005, Dako, Glostrup, Denmark) according to the manufacturer’s instructions. Specific polyclonal primary antibodies were directed against p53 (SAF SP 6874, recombinant human wild type, Sigma, Darmstadt, Germany; dilution 1:4000), ?Np63 (ab735, Abcam, Cambridge, UK; dilution 1:200), TAp73 (NBP2-24737, Novus, Centennial, Colorado, USA; dilution 1:1000), MDM2 (ab3110, Abcam, Cambridge, UK; dilution 1:3000), and γ-H2AX (4411-PC-020, Trevigen, Gaithersburg, MD, USA; dilution 1:250). Antigen retrieval was done with a microwave in a target retrieval solution (cat. no. S1699, Dako, Glostrup, Denmark). The p53 antibody used in this study detected both wild type and mutated p53, with the exception of p53 mutations located between amino acids 20 and 25. Sections were counterstained with Mayer’s hemalum solution and mounted with a cover slide. Primary antibodies were omitted as negative controls.
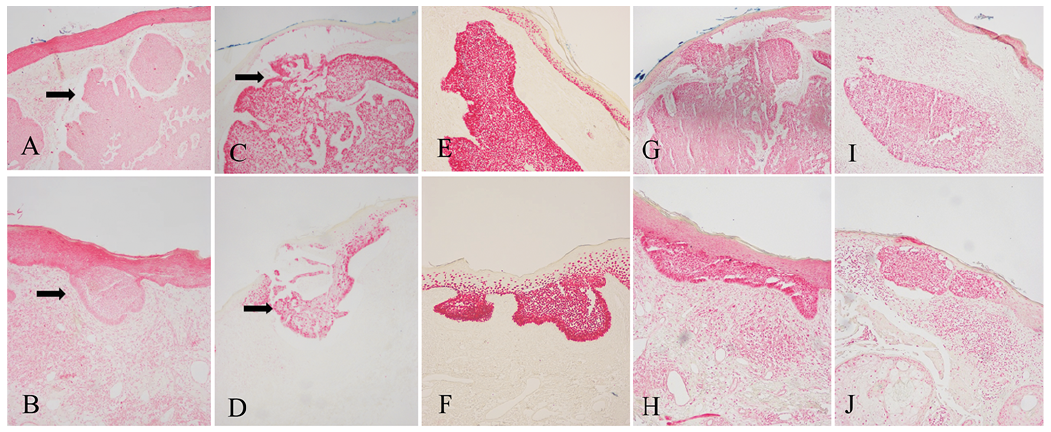
The expression of MDM2, p53, ?Np63, TAp73, and γ-H2AX is shown in Fig. 1 and Table I. Immunoreactivity for p53, ?Np63, TAp73, and γ-H2AX was increased in BCC tumour cells compared with cells of the overlying unaffected epidermis (Fig. 1C–J, respectively). In contrast, MDM2 immunoreactivity was decreased in BCC tumour cells compared with cells of the overlying unaffected epidermis (Fig. 1A, B). In summary, the results of this study showed increased expression of p53, ?Np63, TAp73 and γ-H2AX in BCC, which was associated with decreased expression of MDM2.
Fig. 1. (A) Decreased MDM2 expression compared with the overlying epidermis in nodular basal cell carcinoma (BCC) (black arrow) (×100). (B) Decreased MDM2 expression compared with the neighbouring epidermis in early BCC (black arrow) (×100). (C) Increased p53 expression compared with the overlying epidermis in nodular BCC (×100). (D) Increased p53 expression compared with the neighbouring epidermis in early BCC (×100). (E) Increased ?Np63 expression compared with the overlying epidermis in nodular BCC (×100). (F) Increased ?Np63 expression compared with the neighbouring epidermis in early BCC (×100). (G) Increased TAp73 expression compared with the overlying epidermis in nodular BCC (×100). (H) Increased TAp73 expression compared with the neighbouring epidermis in early BCC (×100). (I) Increased γ-H2AX expression compared with the overlying epidermis in nodular BCC (×100). (J) Increased γ-H2AX expression compared with the neighbouring epidermis in early BCC (×100).
Table I. MDM2, p53, ?Np63, TAp73, and γ-H2AX expression in basal cell carcinoma (BCC)
The tumour suppressor p53 is very important in the pathogenesis of many types of cancers. Although the relevance of p53 mutations for tumorigenesis of BCC is still not completely understood (2, 3), the relatively high rate of p53 UVB signature mutations in BCC implies that UVB may play a role in the subgroup of BCCs harbouring p53 mutations (4). The p63 gene is expressed as 2 isoforms. It is generally assumed that TAp63 is a tumour suppressor gene, while ΔNp63 is an oncogene (5, 6). The p73 gene is also a member of the p53 family, with high structural homology to p53 and capable of transactivating p53 target genes. The p73 gene consists of 2 isoforms, TAp73 (tumour suppressor isoform) and ΔNp73 (oncogenic isoform), with opposing effects, which regulate apoptosis, proliferation, and differentiation (7). It can be speculated whether the increased expression of p53, ΔNp63, TAp73 and γH2AX in the BCCs of the present study are part of the cellular responses to DSBs or other types of DNA damage that may be induced by UV radiation. Moreover, our findings are in agreement with the hypothesis that increased expression of TAp73 may be a cellular mechanism to enhance p53 signalling or to compensate for defective p53 signalling.
MDM2 is an important negative regulator of p53, both as an E3 ubiquitin ligase that recognizes the N-terminal trans-activation domain (TAD) of the p53 tumour suppressor and as an inhibitor of p53 transcriptional activation (8). Millon et al. (9) previously reported decreased MDM2 expression in human head and neck squamous cell carcinomas. In their study, decreased MDM2 expression was significantly associated with advanced tumour stage and poor prognosis. To the best of our knowledge, we show here for the first time decreased MDM2 expression in BCC cells compared with cells of the unaffected overlying epidermis. It is well known that MDM2 promotes the rapid degradation of p53 (8). Therefore, increased expression of p53 in BCC may be, at least in part, due to down-regulated MDM2.
Replication stress, e.g. when UVB-induced DNA lesions collide with replication forks, causes γ-H2AX generation, and the formation of γ-H2AX may serve as an early sensor of DSBs, representing an initial cellular response to UVB-induced DSBs. Moreover, γ-H2AX may also represent a biomarker for the transformation of normal tissue to premalignant and, consequently, to malignant tissues (10). Therefore, it may be speculated whether increased expression of γ-H2AX in the BCCs reported in the present study may be caused by DNA damage induced by UVB or other agents.
In summary, to the best of our knowledge, this is the first paper to show increased expression in BCCs of p53, ?Np63, TAp73 and γ-H2AX associated with decreased expression of MDM2. These findings strongly suggest an important role of p53 signalling for the pathogenesis of BCC. Future studies must analyse whether alterations of p53 signalling represent part of the cellular response to UV radiation-induced DNA damage or if these alterations are caused by other mechanisms.


 Click to show fullsize
Click to show fullsize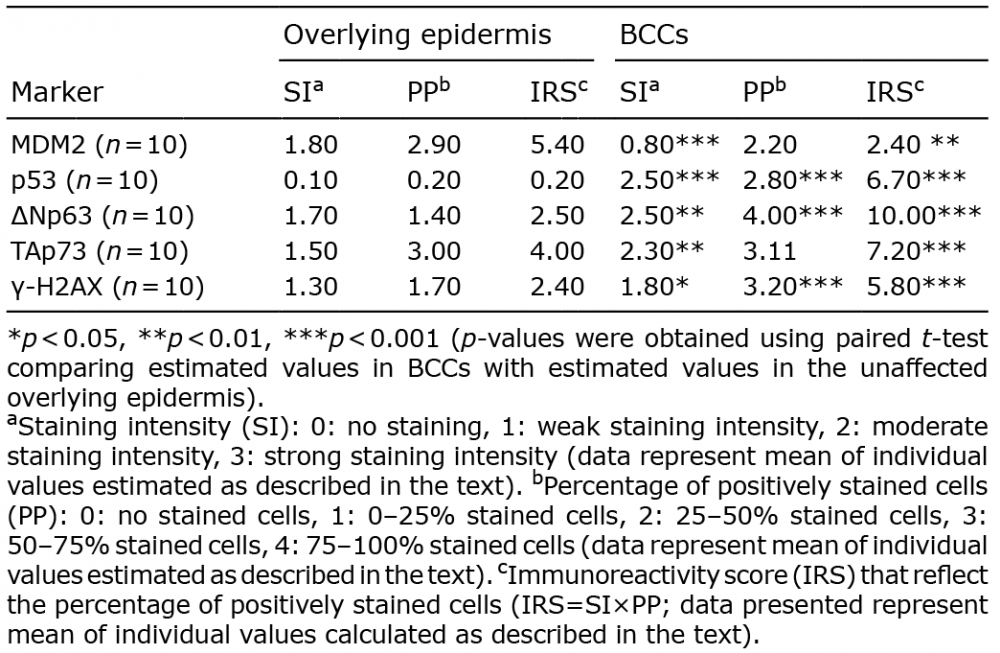 Click to show fullsize
Click to show fullsize