


Claudia Rasche1, Christin Wolfram1, Michael Wahls2 and Margitta Worm1
1Department of Dermatology and Allergy, Charité – Universitätsmedizin Berlin, and 2Department of Otorhinolaryngology, Bundeswehrkrankenhaus Berlin, Germany
Bacterial stimulation plays an important role in modulating the allergic immune response. The aim of this study was to investigate the effects of inactivated probiotic Lactobacillus acidophilus and non-pathogenic Escherichia coli strain Nissle on the phenotype and function of T- and B-cells. Peripheral blood mononuclear cells from patients with grass-pollen allergy (n = 10) and non-allergic patients (n = 19) were co-stimulated with inactivated bacteria and grass-pollen allergen. Expression of CD23, CD80, CD86 and CD69 were analysed, and the intracellular production of interleukin-4 and interferon-γ was measured by direct ex vivo flow cytometry after stimulation. Both bacteria induced a significant up-regulation of CD69 expression on T lymphocytes (p = 0.001). CD23 expression was significantly increased following stimulation with allergen (p = 0.008), but reduced after stimulation with Lactobacillus and significantly reduced with E. coli plus allergen (p = 0.029). CD80 expression was reduced after stimulation with Lactobacillus in the allergic group only (p = 0.021). By contrast, CD86 expression was significantly increased after stimulation with Lactobacillus (p = 0.049) and distinctly increased with E. coli in both groups (p = 0.001). The cytokine patterns of CD69-positive T lymphocytes from allergic patients showed a T-helper2-dominated response after allergen stimulation (interferon-γ/interleukin-4-ratio 0.2), directed into a T-helper1-like response by stimulation with both types of bacteria (interferon-γ/interleukin-4-ratios 1.5–2.0 in both groups). These data show that both types of bacteria modulate the allergic immune response by the alteration of CD23 and co-stimulatory molecule expression. Regarding cytokine production, the data suggest a differential response to both bacteria depending on the atopic state, but a clear promotion of T-helper1-dominated response in allergic donors. Key words: allergy; T-cells; Escherichia coli Nissle 1917; Lactobacillus acidophilus.
(Accepted October 16, 2006.)
Acta Derm Venereol 2007; 87: 305–311.
Claudia Rasche, Charité, Universitätsmedizin Berlin, Department of Dermatology and Allergy, Schumannstr. 20/21, DE-10117 Berlin, Germany. E-mail: margitta.worm@charite.de
New perspectives concerning the prevention of atopic disorders and their treatment are of current interest as these diseases are continuously increasing. Estimates of worldwide prevalence indicate that 20% of the population have asthma and rhinitis, 5% have atopic dermatitis and 3% have food allergy (1).
There are several hypotheses about the causes of this increase, one of the most controversial of these is the so-called “hygiene hypothesis” (2). This associates the increase in allergic diseases with changes in the interactions between people and the microbes in their ecosystem consequent to a “western lifestyle” (3). Extensions of this hypothesis focus on the reduced diversity and magnitude of the “microbial burden” in early life, which is thought to lead to less bacteria-derived maturation signals being encountered during early immune development (4).
Considering this hypothesis, microbial products functioning as immunological stimulators have been assumed to play an important role in the prevention and treatment of allergic diseases (5, 6), but their clinical efficacy has shown diverse results and their usage is preliminary (5). Among these microbial products are the so-called “probiotics”, a group of living or inactivated micro-organisms of potential benefit when ingested (7). The most commonly used probiotics are lactobacilli and bifidobacteria, but enterococci and Escherichia coli have also been proposed as probiotics (5, 8). Previous studies have shown beneficial effects of probiotic micro-organisms, even in adults, caused by promoting the gut barrier function to prevent unfavourable intestinal micro-ecological alterations (9). Research into extensions of the hygiene hypothesis to adults appeared to show that allergic recurrences were improved or prevented in rhinitis patients by eating yoghurt (10), but no effects were seen on birch pollen allergy after oral treatment with Lactobacillus rhamnosus (11).
The present study analysed the influence of chill-inactivated probiotic L. acidophilus and non-pathogenic E. coli Nissle 1917 on the phenotype and function of T- and B-lymphocytes from adult allergic and non-allergic individuals. In order to elucidate the immunomodulatory effects of these bacterial components in humans an in vitro assay was used, followed by direct ex vivo analysis of peripheral lymphocytes, representing systemic bacterial administration and their impact on gut-associated lymphoid tissue lymphocytes.
Previous studies have shown that a T-helper (TH)2-dominated response via the increased production of interleukin (IL)-4 can be arrested by cross-regulatory cytokines such as interferon gamma (IFNγ) (12). Focusing on the allergen-specific immune response, the ability of allergen (A) and of the above-mentioned bacterial components (LC-lactobacilli, EC-E. coli Nissle) to modulate the peripheral TH1/TH2 balance was investigated in adult allergic (a) and non-allergic (na) individuals. In addition, the state of activation and the expression of important co-stimulatory molecules in the presence of the bacteria was analysed by direct ex vivo flow cytometry.
MATERIALS AND METHODS
Patients
The study included 29 age- and sex-matched adult patients from the Department of Otorhinolaryngology (Bundeswehrkrankenhaus, Berlin, Germany), who gave written consent. The study was approved by the local ethics committee. Ten allergic patients suffering from seasonal allergic rhinitis were diagnosed as being sensitized to grass pollen according to skin prick testing (ALK, Hamburg, Germany) and detection of specific IgE (Pharmacia, Freiburg, Germany) to grass pollen (CAP-class ≥2). Nineteen non-allergic, non-grass-pollen sensitized healthy individuals were included as controls. Exclusion criteria were: administration of systemic and local steroids within 14 days prior to the study; the use of antihistamines during the 3 days prior to the study; or current specific immunotherapy or other immunomodulating medication. None of the patients had allergic symptoms at the time of investigation, as the study was performed during the winter season, and none had any other severe diseases.
Cell separation and culture
Heparinized blood samples were taken once from patients and control individuals. Peripheral blood mononuclear cells (PBMCs) were isolated from the heparinized whole blood by Ficoll density gradient centrifugation and washed repeatedly in phosphate buffered saline (PBS) before being resuspended and cultured in RPMI 1640 medium (Sigma, Munich, Germany) supplemented with 10% foetal calf serum, glutamine (2 mM), penicillin (100 U/ml) and streptomycin (100 µg/ml). PBMCs were incubated in flat-bottomed 24-well plates at 37°C in a water-saturated atmosphere containing 5% CO2.
Stimulants
PBMCs were stimulated with grass pollen extract (500 SQ/ml, provided by ALK Scherax, Hamburg, Germany), –80°C chill-inactivated E. coli Nissle (Mutaflor, Ardeypharm, Herdecke, Germany) at a 1:1 ratio, or L. acidophilus (Paidoflor, Ardeypharm) at a 2:1 ratio (13). The grass pollen extract additionally contained anti-CD28 (0.5 µg/ml, BD PharMingen, Heidelberg, Germany) for co-stimulation. As a positive control, PBMCs were incubated with staphylococcal enterotoxin B (SEB) (1 µg/ml, Sigma). The negative control contained unstimulated cells. PBMCs were collected after 18 h for surface staining and after 16 h for intracellular staining. Grass pollen extract was analysed with regard to endotoxin content, measured by the chromogenic Limulus amebocyte lysate assay (Bio Whittaker) and showed 222 pg/ml in the allergen dilution of 100,000 SQ/ml. Accordingly, 500 SQ/ml of allergen extract contains a negligible amount of endotoxin, which has no modulating effect.
Flow cytometry
The expression of surface CD20, CD23, CD80, CD86, CD4 and CD69, and of intracellular IL-4 and IFNγ were assayed by flow cytometry (all antibodies were "mouse-anti-human" from BD/PharMingen). PBMCs (1×106) were incubated with the specific antibodies and the appropriate isotype controls (BD/PharMingen) for 30 min at 4°C in 100 µl staining buffer (2% bovine serum albumin in PBS, 0.002% sodium azide) following a 10-min-blocking with 5 µl Beriglobin (Aventis-Behring, Marburg, Germany). Cells for intracellular staining were additionally incubated with Brefeldin A (1 µl/ml, Sigma) for 3 h after 13 h of incubation, and permeabilized with 500 µl FACS-permeabilizing solution (BD Bioscience, Heidelberg, Germany) before staining. Finally, the cells were washed and fixed in PBS with 2% paraformaldehyde.
Flow cytometry of 4-fold labelled cells was performed with a FACScalibur (Becton Dickenson, Heidelberg, Germany) flow cytometer and evaluated using Cell Quest software (Becton Dickenson). Lymphocytes were gated by anti-CD20 or anti-CD4 fluorescence, the percentage of the double-positive cells after deduction of the isotype control values was determined.
Preceding analysis
Dose-response experiments were initially carried out using increasing bacterial concentrations (i.e. 0.25×106, 0.5×106, 1×106 and 2×106 E. coli / L. acidophilus per 1×106 cells) for stimulation. Subsequently, a time course analysis covering 6, 18, 24 and 48 h of stimulation was performed, using the determined bacterial concentration. PBMCs were stained with CD4 and CD69 Allophycocyanin, respectively, CD20 and CD23, CD54 or CD86 to analyse their level of activation (data not shown). Mitochondrial activity in response to all the stimulants used was analysed using 10 μl WST-1 reagent (4-(3-(4-iodophenyl)-2-(4-nitrophenyl)-2H-5-tetrazolio)-1,3-benzene disulphonate, Boehringer Mannheim) per 100 μl cell-stimulant preparation (as described above) and measured half-hourly for 4 h by spectophotometric quantification (OD 450 nm) after 18 h of stimulation.
Statistical analysis
Analysis of the statistical significance of within-group changes was carried out using Brunner’s analysis and the Wilcoxon test for non-parametric data. Descriptive statistical analysis, table calculations and diagrams were produced using Excel 2000 and SPSS 11.0. The values in Table I represent the mean ± standard error of the mean or ratio of means.
Table I. Intracellular measurement of cytokine production by CD4-positive T lymphocytes in allergic (a) and non-allergic (na) individuals. Expression of interleukin (IL)-4 and interferon (IFN)γ on CD4+ T-lymphocytes, determined by flow cytometry in unstimulated cells (U) and following incubation with grass pollen allergen (A), E. coli Nissle (EC) and L. acidophilus (LC). Each value represents the mean ± SEM as a percentage of CD4+ T-lymphocytes
| CD4+ | ||||
| IL-4 | IFNγ | |||
| a | na | a | na | |
| U | 0.27 ± 0.14 | 0.55 ± 0.09 | 0.10 ± 0.08 | 0.31 ±0.05 |
| A | 0.34 ± 0.18 | 0.66 ± 0.08 | 0.16 ± 0.08 | 0.43 ± 0.07 |
| LC | 0.65 ± 0.28* | 0.97 ± 0.23* | 0.48 ± 0.18* | 0.87 ± 0.15* |
| LC+A | 0.91 ± 0.32** | 1.08 ± 0.22** | 0.49 ± 0.18** | 0.74 ± 0.19* |
| EC | 0.17 ± 0.14 | 0.88 ± 0.19 * | 0.14 ±0.12 | 0.70 ± 0.20* |
| EC+A | 0.37 ± 0.19 | 0.91± 0.30** | 0.57 ± 0.21*** | 0.80 ± 0.25** |
*p <0.05 compared with U; **p <0.05 compared with U, A; ***p <0.05 compared with U, A, EC.
RESULTS
Mitochondrial activity assay
With respect to cell viability, the addition of bacteria caused a reduced mitochondrial activity strongest after stimulation with L. acidophilus (ODLC 0.18; ODEC 0.19) in comparison with SEB (OD 0.4) and allergen-stimulated (ODA 0.29) as well as unstimulated cells (ODU 0.28). Incubation of bacteria alone did not show any effect on the mitochondrial activity measured.
Increased T-cell activation by microbacterial stimulation
First, the T-lymphocytes were investigated with regard to the state of activation via surface CD69 expression (14). A significant up-regulation of CD69 expression was detected following allergenic (median: Aa 3.9%, pa = 0.004; Ana 2.9%, pna = 0.001) and bacterial stimulation with L. acidophilus in both groups (median: LCa 5.0%; LC+Aa 5.7%, pa = 0.001; LCna 3.7%; LC+Ana 4.4%, pna = 0.001) in comparison with basal CD69 expression (median: Ua 2.1%; Una 2.0%). In the presence of E. coli, CD69 expression showed a distinct up-regulation, which was highly significant with or without the addition of allergen (median: ECa 29.2%; EC+Aa 36.0%, pa = 0.001; ECna 23.8%; EC+Ana 29.5%, pna = 0.001).
Triggering of TH1-dominated T-cell responses by L. acidophilus, lactobacilli and E. coli
As the balance of TH1/TH2 response plays a central role for the development of allergic disease (15), intracellular IL-4 and IFNγ production was measured on CD4-positive T-lymphocytes by intracellular staining (Fig. 1). Stimulation with grass pollen extract led to an increase in IL-4 production (median: Ua 0.27%; Aa 0.34%) and, to a lesser extent in IFNγ production in the allergic group (median: Ua 0.1%; Aa 0.16%).
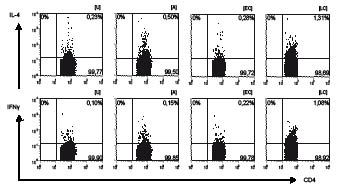
Fig. 1. Representative example of the intracellular production of IL-4 and IFNγ in the presence of E. coli Nissle 1917 (EC) and L. acidophilus (LC) and combined stimulation with allergen (EC+A, LC+A) compared with unstimulated T-cells (U) and allergen (A). Scatter plots, showing intracellular flow cytometry of cytokines in peripheral blood CD4-positive T lymphocytes in allergic individuals following 16 h stimulation (see Materials and Methods).
Analysis of the IFNγ/IL-4-ratio of allergen specific T-lymphocytes is presented in Fig. 2 and shows a TH2-dominated response after allergen exposure, especially in the allergic group (CD69+ IFNγ/IL-4-ratioA 0.2a; 0.75na). In contrast to these findings, TH1-dominated responses were observed following the incubation with L. acidophilus in both groups (CD69+ IFNγ/IL-4-ratioLC 2.0a; 1.5na). However, stimulation with lactobacilli plus allergen resulted in a TH2-like response in allergic and non-allergic individuals (CD69+ IFNγ/IL-4-ratioLC+A 0.67a; 0.84na). As seen in Table I, this may be due to a significant increase in IL-4 production (median: LC+Aa 0.91%; LC+Ana 1.08%, p = 0.041), whereas IFNγ production remained basically unchanged in the allergic group (median: LCa 0.48%; LC+Aa 0.49%; LCna 0.87%; LC+Ana 0.74%).
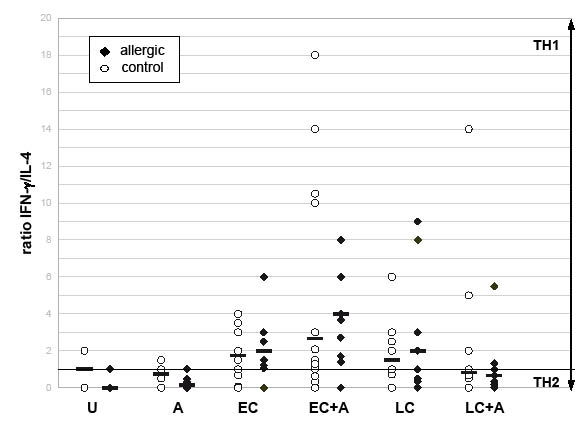
Fig. 2. Induction of TH1-dominated immune response in activated CD4-positive T-lymphocytes following stimulation with non-pathogenic E. coli Nissle (EC) and L. acidophilus (LC) in comparison with unstimulated cells (U) and stimulation with allergen (A). Ratios calculated from the percentages of CD69-IL-4 and CD69-IFNγ double-labelled CD4-positive T-lymphocytes, na = 7, nna = 13. A ratio less than 1 means a predominance of IL4 compared with IFNγ, representing a TH2 response, and vice versa.
A significant increase in IFNγ production was present after stimulation with E. coli and allergen (median: EC+Aa 0.57%, pa = 0.007; EC+Ana 0.80%, p = 0.001), whereas single E. coli-stimulation caused a strong up-regulation of IFNγ in non-allergic individuals only (median: ECa 0.14%; ECna 0.70%; pna = 0.001, see Table I). Finally, E. coli also had little effect on the production of IL-4 in the allergic group. Non-allergic subjects showed a significant increase (median: ECa 0.17%; ECna 0.88%; pna = 0.001). As shown in Fig. 2, the IFNγ/IL-4-ratio of allergen-specific T-lymphocytes reflects these findings of a TH1-dominated response following exposure to E. coli (CD69+ IFNγ/IL-4-ratioEC 2.0a; 1.75na) as well as E. coli and allergen (CD69+ IFNγ/IL-4-ratioEC+A 4.0a; 2.66na).
E. coli stimulation enhanced CD80 and CD86 expression
The importance of co-stimulatory molecules, especially the up-regulation of CD86 but not CD80 following allergenic or IL-4 stimulation in the development of TH2-cells in patients with atopic diseases has been described repeatedly (16, 17). Our results show that grass pollen extract induced a small, but significant, increase in CD86 expression only in the allergic subjects (median: Ua 4.26%; Aa 5.6%; p = 0.025) and had no impact on the expression of CD80 on B-lymphocytes (median: Ua 10.6%; Aa 10.4%).
We then investigated the influence of the above-mentioned bacteria on the expression of co-stimulatory molecules. As shown in Fig. 3, stimulation with L. acidophilus reduced the percentage of B-cells positive for CD80 in the allergic group (median: LCa 7.4%, p = 0.021). E. coli caused an up-regulation of CD80 in both groups, which was statistically significant only in the non-allergic subjects (median: ECa 13%; ECna 16%; pna = 0.001) and similarly after stimulation with E. coli and allergen (median: EC+Aa 13%; EC+Ana 16%, p = 0.001).
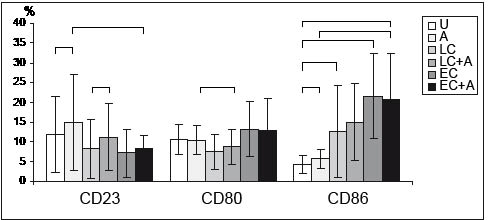
Fig. 3. CD23 and co-stimulatory molecule expression upon stimulation with bacteria. Percentage of CD23-, CD80- and CD86-positive B-cells in allergic individuals. Flow cytometry was performed following 18 h of stimulation. Mean values ± SEM (brackets indicate p <0.05). U: unstimulated (n = 8); A: allergen (n = 8); LC: lactobacilli (n = 7); EC: E. coli Nissle (n = 6); LC+A (n=7); EC+A(n=6).
As shown in Fig. 4, CD86-expression on B lymphocytes following bacterial stimulation showed a significant increase in both groups (median: ECa 21%; EC+Aa 21%; ECna 20.7%; EC+Ana 24.8%, pa = 0.007, pna = 0.001; LCa 12.6%; LCna 9.9%, pa = 0.049, pna = 0.001), except for the increase following L. acidophilus plus allergen, which did not reach statistical significance (median: LC+Aa 15%; LC+Ana 6.5%).
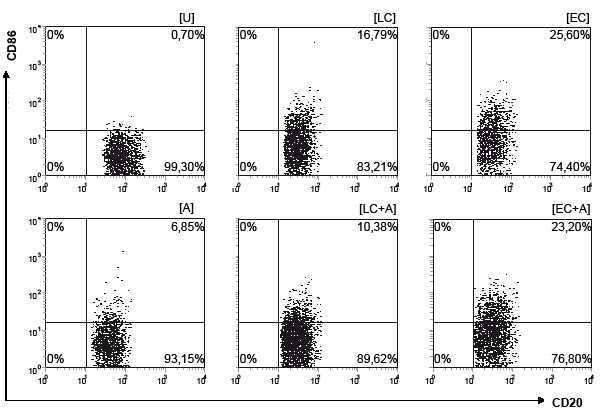
Fig. 4. Increase in CD86 expression following stimulation with grass pollen allergen (A), E. coli Nissle 1917 (EC) and L. acidophilus (LC) in comparison with unstimulated cells (U). Scatter plots of flow cytometry of peripheral blood CD20-positive B-lymphocytes in allergic individuals following 18 h stimulation (see Materials and Methods).
Modulation of CD23 expression by inactivated bacteria
CD23, the IL-4-induced and IFNγ-modulated low-affinity receptor for IgE, plays a crucial role in regulating T- and B-cell interactions within the allergic response (18). Therefore we analysed the effect of bacterial and allergenic stimuli on the expression of CD23 on peripheral B-lymphocytes in allergic and non-allergic subjects. We observed higher basal expression and significantly higher up-regulation of CD23 after exposure to allergen within the allergic group (median: Ua 11.9%; Aa 15.0%, p = 0.012) than the non-allergic control (median: Una 5.1%; Ana 5.4%). E. coli reduced the expression of CD23 compared with allergenic stimulation (median: ECa 7.1%; ECna 6.0%), which appeared to be significant after co-stimulation with E. coli and allergen in the allergic group only (median: EC+Aa 8.2%; EC+Ana 5.7%, pa = 0.029).
Incubation with L. acidophilus led to decreased expression of surface CD23 in the allergic group in particular, but this did not reach statistical significance (median: LCa 8.3%). The addition of allergen increased the percentage of CD23-positive B-cells significantly compared with L. acidophilus in both groups (median: LC+Aa 11.2%; LC+Ana 9.3%; pa = 0.001, pna = 0.03).
DISCUSSION
This study investigated the immunomodulatory effects of inactivated probiotic L. acidophilus and non-pathogenic E. coli in an ex vivo model on peripheral lymphocytes from sensitized allergic individuals and healthy controls.
The results show that stimulation with allergen caused a TH2-like response with enhanced IL-4 production in both groups and up-regulation of CD86 and CD23 on B-lymphocytes in allergic individuals. This immune response was profoundly modulated by the bacterial components L. acidophilus and E. coli. Single bacterial stimulation resulted in a TH1-dominated response in both groups, as reported in previous clinical studies (5). Furthermore, the expression of CD23, but also the co-stimulatory molecules CD80 and CD86 were modulated by both L. acidophilus and E. coli.
The shift in the TH1/TH2-balance measured via IL-4 and IFNγ following stimulation with non-pathogenic bacteria demonstrates the possible therapeutic potency especially of non-pathogenic E. coli Nissle. Although some studies indicate that human cytokine activities fail to exhibit an exclusively pro-TH1 or -TH2 pattern (19), our results show rather a reduction in one pathway.
Moreover, allergic patients showed decreased CD23 expression in the presence of bacteria, especially E. coli. These observations are closely related to the modulating effects of IL-4 and IFNγ on the expression of CD23. As previous studies describe increased CD23 expression caused by IL-4 and a decreased CD23 expression caused by IFNγ, the above-mentioned cytokine patterns following bacterial stimulation explain our findings (18). The in vivo relevance has been shown, but could not be confirmed clinically (20, 21). Increased CD23 expression in patients suffering from atopic dermatitis and allergic rhinitis has been shown previously by our group (22). Furthermore, former studies show that IFNγ inhibits CD23 mRNA and sCD23 production on human B-cells (18). sCD23, which was not measured in this study, has cytokine properties alone, but is also able to elicit strong inflammatory action by stimulating the production of TNFα and IL-1β and co-stimulates IL-2-induced INFγ production (23). Decreased shedding of sCD23 results in decreased IgE production (24).
In addition, the co-stimulatory molecules CD80 and CD86 were modulated by both types of bacteria. Non-pathogenic E. coli resulted in up-regulation of CD80 and CD86, indicating a non-specific stimulation of the B-cell system. This was consistent with with an increased production of specific antibodies and proliferative activity, as described previously by Cukrowska et al. (25). By contrast, incubation with probiotic L. acidophilus led to inhibition of CD80 but up-regulation of CD86 especially in allergic individuals. Thus, our findings may support the hypothesis of CD80 functioning as a more neutral differentiation signal instead of a TH1-maintaining one and – also explaining the E. coli-derived results – of both molecules directing the development of TH1 and TH2 in a less contrasting pattern (17, 26).
As it is shown that T-cell activation does not necessarily need antigen presenting cell signalling for activation (27), differences between the bacteria may be based on toll-like receptor (TLR)-mediated regulation of pathogen-related responses, since there are different cascades of activation mechanisms of Gram-positive and Gram-negative bacteria (28, 29). Without focusing on TLR-mediated cascades, the basic activity of lymphocytes plays a crucial role in the distinct TH1-like responses on bacterial stimulation in the presence of allergen, especially the allergic group. If one assumes preventive or even maturating effects of L. acidophilus and even non-pathogenic E. coli beyond childhood, these may be also explained via the TLR model. As the hygiene hypothesis focuses only on the maturation of the immune system towards TH1 phenotype in childhood (30), TLR pathways may also be involved in the maturation of the normal adult immune system. It remains unclear whether the above-mentioned TH1-responses following stimulation with probiotic and apathogenic bacteria as seen in our study show prolonged effects on driving the maturation of the adult immune system. However, the current study does not lead to precise conclusions about the comparative potency of both types of bacteria.
The strong TH1-like responses following bacterial stimulation also seen in the non-allergic controls suggest that priming signals may exist and consequently prevent over-expression of TH2-dominated responses. These effects might play a role not only in childhood (31). Previous studies using murine models indicate that such immunomodulatory effects are derived from long-term administration of mutated bacterial agents. Such effects may also result in long-term down-regulation of hypersensitivity reactions secondary to antigen-specific TH1/TH2 imbalances (32). As it is even hypothesized that TH1- and TH2-mediated conditions might frequently coexist and that the contrasting TH1/TH2-model might be an oversimplification (33), we assume, that long-term immunomodulating is possible beyond priming in infancy.
Another interesting observation is the shift of the allergen-specific TH2 response towards a TH1-like response following stimulation with E. coli Nissle plus allergen. This effect was not seen with L. acidophilus. Whether these observations reflect the clinical findings of a poor reduction in symptoms after oral L. acidophilus treatment requires further investigation (34). On the other hand, oral E. coli application has been reported to reduce the incidence of IgE-dependent allergies, namely respiratory and skin manifestations, in children when administered after birth (6, 34).
In summary, our observations indicate that inactivated non-pathogenic E. coli and probiotic L. acidophilus have immunomodulating capacities, as we determined profound changes in the TH balance. Clinical studies also indicate the applicability of probiotic agents for preventive use (35, 36).
The promotion of a TH1-dominated immune response by long-term application of bacteria, and especially E. coli, may be critical, as autoimmune diseases are characterized by a TH1-dominated immune response with high levels of TH1-type cytokines, e.g. IFNγ. On the other hand, it has recently been shown in a clinical trial that oral administration of E. coli Nissle maintains the remission of ulcerative colitis, an autoimmune disease of the gut (37).
E. coli Nissle, but not lactobacilli, were shown to induce high levels of IL-10 and IL-12, which might be due to the content of lipopolysaccharides in Gram-negative bacteria only. Therefore IL-12-promoted induction of IFNγ may be the cause of the differences in modulation of TH1/TH2 balance seen when comparing the types of bacteria. Finally, changes in the expression of surface molecules, such as CD86 and CD23, might also originate from this pro-inflammatory immune response (38).
We conclude that probiotic L. acidophilus and non-pathogenic E. coli Nissle exhibit immunomodulatory effects on lymphocytes by influencing the cytokine pattern, indicating a potential for preventive and therapeutic efficacy in the treatment of TH2-dominated allergic diseases. E. coli in particular increases the production of IFNγ under allergen challenge, which might also result in reduced symptoms in allergic individuals. However, in order to elucidate the role of non-pathogenic E. coli Nissle in modulating allergic symptoms and its mechanism of action, further investigation are needed.
REFERENCES
1. Baena-Cagnani CE, Serra H, Teijeiro A, Croce JS. Prevention of allergy and asthma. Clin Exp All Rev 2003; 3: 51–57.
2. Strachan DP. Hay fever, hygiene, and household size. BMJ 1989; 299: 1259–1260.
3. Matricardi PM, Bonini S. High microbial turnover rate preventing atopy: a solution to inconsistencies impinging on the Hygiene hypothesis? Clin Exp Allergy 2000; 30: 1506–1510.
4. Prescott SL. Allergy: the price we pay for cleaner living? Ann Allergy Asthma Immunol 2003; 90: 64–70.
5. Matricardi PM, Bjorksten B, Bonini S, Bousquet J, Djukanovic R, Dreborg S, et al. Microbial products in allergy prevention and therapy. Allergy 2003; 58: 461–471.
6. Lodinova-Zadnikova R, Cukrowska B, Tlaskalova-Hogenova H. Oral administration of probiotic Escherichia coli after birth reduces frequency of allergies and repeated infections later in life (after 10 and 20 years). Int Arch Allergy Immunol 2003; 131: 209–211.
7. Schrezenmeir J, de Vrese M. Probiotics, prebiotics, and synbiotics – approaching a definition. Am J Clin Nutr 2001; 73: S361–S364.
8. Jansen GJ, Wildeboer-Veloo AC, van der Waaij D, Degener JE. Escherichia coli as a probiotic? Infection 1998; 26: 232–233.
9. Bongaerts GP, Severijnen RS. Preventive and curative effects of probiotics in atopic patients. Med Hypotheses. 2005; 64: 1089–1092.
10. Aldinucci C, Bellussi L, Monciatti G, Passali GC, Salerni L, Passali D, et al. Effects of dietary yoghurt on immunological and clinical parameters of rhinopathic patients. Eur J Clin Nutr 2002; 56: 1155–1161.
11. Helin T, Haahtela S, Haahtela T. No effect of oral treatment with an intestinal bacterial strain, Lactobacillus rhamnosus (ATCC 53103), on birch-pollen allergy: a placebo-controlled double-blind study. Allergy 2002; 57: 243–246.
12. Cross ML, Stevenson LM, Gill HS. Anti-allergy properties of fermented foods: an important immunoregulatory mechanism of lactic acid bacteria? Int Immunopharmacol 2001; 1: 891–901.
13. Baati L, Fabre-Gea C, Auriol D, Blanc PJ. Study of the cryotolerance of Lactobacillus acidophilus: effect of culture and freezing conditions on the viability and cellular protein levels. Int J Food Microbiol 2000; 59: 241–247.
14. Testi R, Phillips JH, Lanier LL. T cell activation via Leu-23 (CD69). J Immunol 1989; 143: 1123–1128.
15. Martinez FD. The coming-of-age of the hygiene hypothesis. Respir Res 2001; 2: 129–132.
16. Nakada M, Nishizaki K, Yoshino T, Okano M, Masuda Y, Ohta N, et al. CD86 (B7-2) antigen on B cells from atopic patients shows selective, antigen-specific upregulation. Allergy 1998; 53: 527–531.
17. Hofer MF, Jirapongsananuruk O, Trumble AE, Leung DY. Upregulation of B7.2, but not B7.1, on B cells from patients with allergic asthma. J Allergy Clin Immunol 1998; 101: 96–102.
18. Corominas M, Mestre M, Bas J, Buendia E. Distinct modulation by interferon-gamma (IFN-gamma) of CD23 expression on B and T lymphocytes of atopic subjects. Clin Exp Immunol 1998; 112: 276–280.
19. Kidd P. Th1/Th2 balance: the hypothesis, its limitations, and implications for health and disease. Altern Med Rev 2003; 8: 223–246.
20. Hanifin JM, Schneider LC, Leung DY, Ellis CN, Jaffe HS, Izu AE, et al. Recombinant interferon gamma therapy for atopic dermatitis. J Am Acad Dermatol 1993; 28: 189–197.
21. Reinhold U, Kukel S, Brzoska J, Kreysel HW. Systemic interferon gamma treatment in severe atopic dermatitis. J Am Acad Dermatol 1993; 29: 58–63.
22. Frotscher B, Anton K, Worm M. Inhibition of IgE production by the imidazoquinoline resiquimod in nonallergic and allergic donors. J Invest Dermatol 2002; 119: 1059–1064.
23. Daniels BB, Askew SL, van de Venter M, Oosthuizen V. Production of biologically active recombinant human soluble CD23 and its effect on PBMCs isolated from hyper-IgE blood. Cell Immunol 2005; 234: 146–153.
24. Wheeler DJ, Parveen S, Pollock K, Williams RJ. Inhibition of sCD23 and immunoglobulin E release from human B cells by a metalloproteinase inhibitor, GI 129471. Immunology 1998; 95: 105–110.
25. Cukrowska B, LodInova-ZadnIkova R, Enders C, Sonnenborn U, Schulze J, Tlaskalova-Hogenova H. Specific proliferative and antibody responses of premature infants to intestinal colonization with nonpathogenic probiotic E. coli strain Nissle 1917. Scand J Immunol 2002; 55: 204–209.
26. Thompson CB. Distinct roles for the costimulatory ligands B7-1 and B7-2 in T helper cell differentiation? Cell 1995; 81: 979–982.
27. Zanin-Zhorov A, Cahalon L, Tal G, Margalit R, Lider O, Cohen IR. Heat shock protein 60 enhances CD4 CD25 regulatory T cell function via innate TLR2 signaling. J Clin Invest 2006; 116: 2022–2032. Epub 2006 Jun 2028.
28. Muzio M, Mantovani A. The Toll receptor family. Allergy 2001; 56: 103–108.
29. Sabroe I, Parker LC, Wilson AG, Whyte MK, Dower SK. Toll-like receptors: their role in allergy and non-allergic inflammatory disease. Clin Exp Allergy 2002; 32: 984–989.
30. von Mutius E, Braun-Fahrlander C, Schierl R, Riedler J, Ehlermann S, Maisch S, et al. Exposure to endotoxin or other bacterial components might protect against the development of atopy. Clin Exp Allergy 2000; 30: 1230–1234.
31. Cross ML, Gill HS. Can immunoregulatory lactic acid bacteria be used as dietary supplements to limit allergies? Int Arch Allergy Immunol 2001; 125: 112–119.
32. Li XM, Srivastava K, Grishin A, Huang CK, Schofield B, Burks W, et al. Persistent protective effect of heat-killed Escherichia coli producing “engineered,” recombinant peanut proteins in a murine model of peanut allergy. J Allergy Clin Immunol 2003; 112: 159–167.
33. Sheikh A, Smeeth L, Hubbard R. There is no evidence of an inverse relationship between TH2-mediated atopy and TH1-mediated autoimmune disorders: Lack of support for the hygiene hypothesis. J Allergy Clin Immunol 2003; 111: 131–135.
34. Wheeler JG, Shema SJ, Bogle ML, Shirrell MA, Burks AW, Pittler A, et al. Immune and clinical impact of Lactobacillus acidophilus on asthma. Ann Allergy Asthma Immunol 1997; 79: 229–233.
35. Isolauri E, Arvola T, Sutas Y, Moilanen E, Salminen S. Probiotics in the management of atopic eczema. Clin Exp Allergy 2000; 30: 1604–1610.
36. Kalliomäki M, Salminen S, Arvilommi H, Kero P, Koskinen P, Isolauri E. Probiotics in primary prevention of atopic disease: a randomised placebo-controlled trial. Lancet 2001; 357: 1076–1079.
37. Kruis W, Fric P, Pokrotnieks J, Lukas M, Fixa B, Kascak M, et al. Maintaining remission of ulcerative colitis with the probiotic Escherichia coli Nissle 1917 is as effective as with standard mesalazine. Gut 2004; 53: 1617–1623.
38. Cross ML, Ganner A, Teilab D, Fray LM. Patterns of cytokine induction by gram-positive and gram-negative probiotic bacteria. FEMS Immunol Med Microbiol 2004; 42: 173–180.

