


Departments of 1Dermatology, Venereology and Allergology, 2Molecular Medicine, and 3Plastic Surgery, Medical University of Gdansk, Gdansk, Poland
Changes in the expression of cornified envelope (CE) proteins are thought to affect the development and course of atopic dermatitis (AD). The aim of this study was to examine the expression level of CE proteins in order to identify new molecular markers of the AD phenotype. Expression levels of CE proteins were evaluated in the skin of patients with AD (38 biopsies) and healthy subjects (26 biopsies). Levels of FLG, FLG2 and SPRR3 mRNAs and proteins were reduced in AD skin. Levels of LELP-1 and SPRR1A transcripts and proteins were significantly increased in AD skin. SPRR3v2 mRNA level in non-lesional AD skin correlated with severity of AD, and SPRR3 protein level in non-lesional AD skin correlated inversely with pruritus. FLG protein level in AD skin correlated inversely with severity of AD. These results point to SPRR3 as an important factor in AD and itch.
Key words: atopic dermatitis; gene expression; cornified envelope proteins; SPRR3.
Accepted Jun 1, 2016; Epub ahead of print Jun 15, 2016
Acta Derm Venereol 2017; 97: 36–41.
Corr: Magdalena Trzeciak, Department of Dermatology, Venereology and Allergology, Medical University of Gdansk, ul. Debinki 7, 80-211 Gdansk, Poland. E-mail: mtrzeciak@gumed.ed
Atopic dermatitis (AD) is a chronic inflammatory skin disease, which is diverse in terms of onset, natural history, clinical manifestation, concomitant allergies and psychological involvement (1). Complex genetic and environmental factors are involved in the development of AD (2). Skin abnormalities are found in the stratum corneum (SC) and tight junction alterations (3), affecting the lamellar body secretory system, lipid structures, and function and formation of cornified envelope (CE) proteins (3–5). The genes encoding CE proteins are clustered in the epidermal differentiation complex (EDC), located on chromosome 1q21.
FLG and its mutations play an important role in AD skin barrier defects and are the best-known risk factors for development of AD (6). Up to 50% of European subjects with AD bear certain FLG mutation, but these mutations are also present in ichthyosis vulgaris and in approximately 9% of the European population, without concomitant inflammation (7). Moreover, there are ethnic and regional differences in FLG mutations (8, 9). However, impaired skin barrier function has also been observed, as shown in patients with AD without FLG mutations (10). Immunological factors, including several TH2 cytokines, as well as environmental factors, reduce expression of filaggrin (11).
It is assumed that, in addition to FLG alterations, changes in other CE proteins may also influence the skin barrier defect and modulate AD course and risk. However, our knowledge of CE proteins other than FLG in the pathogenesis of AD is limited. Thus, the aim of this study was to search for factors associated with AD course and development, focusing on the determination of the expression levels of genes encoding the CE proteins, namely: the late cornified envelope-like proline-rich protein-1 (LELP-1), small proline-rich proteins (SPRR1A, SPRR1B, SPRR3), and filaggrin (FLG, FLG2) in lesional, and non-lesional skin of patients with AD.
Patients with AD were recruited in the Department of Dermatology, Venereology and Allergology on the basis of diagnostic criteria of Hanifin & Rajka (12). The AD group (19 patients) comprised 7 males (36.8%) and 12 females (63.2%), with a mean ± standard deviation (SD) age of 32.6 ± 12.5 years (range 14–59 years). The mean Severity Score of Atopic Dermatitis (SCORAD) (13, 14) was 62.17 ± 14.18 (range 35–86). According to current SCORAD severity classification recommendations the study population may be considered severe. Mean ± SD pruritus was 7.10 ± 1.85 points; mean ± SD IgE level was 2,376 ± 2,021 kU/l. Mean ± SD age at disease onset was 7.07 ± 9.44 years. Patients on immunosuppressive treatment or other immunotherapies were excluded from the study. The control group comprised 26 healthy, ethnically-matched volunteers with no medical history of allergies, immunological diseases or malignancies. The control group comprised 12 males (46.2%) and 14 females (53.8%), mean ± SD age 38.6 ± 12.1 years (range 22–61 years), and mean ± SD IgE level was 35.26 ± 37.62 kU/l.
The study was conducted with the consent (NKEBN/486/2011) of the local ethics committee (Independent Bioethics Commission for Research at Medical University of Gdansk). Written consent was obtained from all patients prior to enrollment in the study.
For the study 38 samples of arm skin (2–4-mm punch biopsies of 3-mm depth) were taken from patients with AD (19 from lesional and 19 from non-lesional AD skin), and 26 skin samples (controls) were obtained from healthy subjects. Because the quantity of available material in skin samples obtained from 2-mm punch biopsies was limited, some samples (13 samples of non-lesional AD skin, 13 of lesional AD skin, and 13 of control skin) were used for gene expression analysis only, and other (4-mm punch biopsies) samples (6 of non-lesional AD skin, 6 of lesional AD skin, and 14 of control skin) were used for gene expression determination and protein level measurement. The non-lesional skin biopsy was performed at (at least) a 10-cm distance from AD skin lesions. Immediately after biopsy the skin samples were placed in RNAlater solution (Qiagen, Hilden, Germany) and stored at –20°C.
Total serum IgE levels were estimated by fluorescent enzyme immunoassay using the Uni-CAP 100 System (Phadia, Sweden) according to the manufacturer’s instructions.
The SCORAD scale helps to determine the severity of the diseases: mild AD correlated with a SCORAD < 25 points, and severe AD with a SCORAD > 50 points (13, 14). Visual analogue scale (VAS)/numeric rating scale (NRS) was employed to estimate the level of pruritus (0–10 points).
Total RNA was isolated using standard methods. mRNA levels were analysed by real-time PCR (RT-PCR) with TaqMan primer-probe sets using the Path-ID Multiplex One-Step RT-PCR kit (Path-ID™ Multiplex One-Step RT-PCR Kit, Applied Biosystems, Foster City, CA, USA). The reference transcript (ACTB or TBP or G6PD) was used as an internal standard and was amplified together with each target gene transcript in the same well using primers and probes (Table SI).
The level of each analysed transcript was normalized to that of the appropriate reference transcript. Data analysis was performed using LightCycler 480 II software (Roche Diagnostics International Ltd, Rotkreuz, Switzerland).
Skin specimens were weighed, rinsed with cold phosphate-buffered saline (PBS) and homogenized in PBS containing 0.5% Igepol CA-630 (1:2) on ice. The homogenates were subjected to 2 freeze-thaw cycles to break the cell membranes, and the homogenates were centrifuged for 5 min at 5,000 g. The supernatants obtained were used for protein determination by enzyme-linked immunosorbent assay (ELISA) kit according to the manufacturer’s protocol (Antibody-Protein ELISA kit, MyBioSource, Inc., San Diego, CA, USA). The quantity of investigated protein in the sample was interpolated from the standard curve and corrected for dilution. Protein names were abbreviated according to HUGO Gene Nomenclature Committee guidelines.
Data from the inquiry prepared for this study were statistically analysed using Excel 2003 (Microsoft Corp., Redmond, WA, USA) and STATISTICA (Version 8.0: StatSoft, Tulsa, OK, USA). Expression data were compared by Mann-Whitney test. For the comparison between lesional and non-lesional skin from the same patient the Wilcoxon matched-pairs test was used. Associations between the 2 variables were assessed based on Spearman’s rank correlation coefficient (Rs). p < 0.05 was regarded as statistically significant.
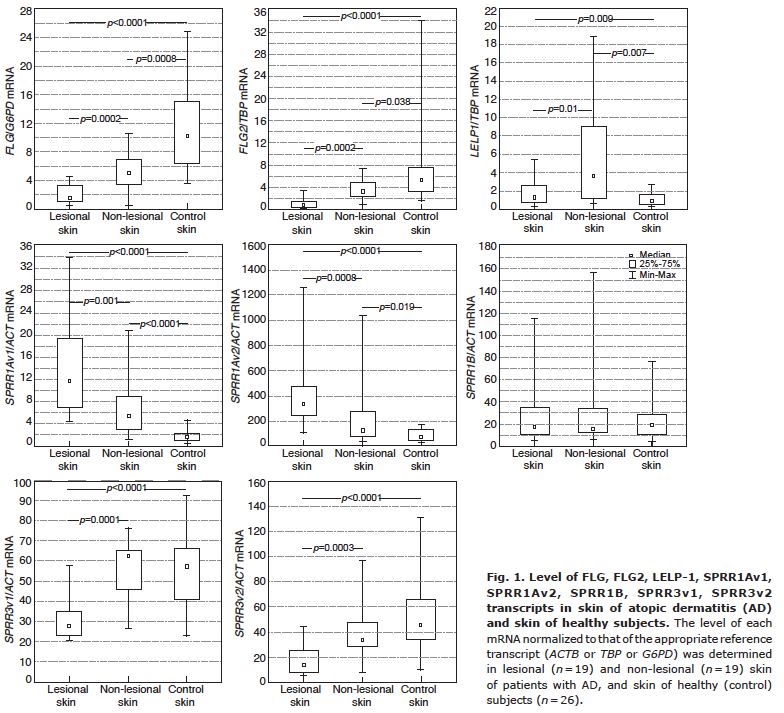
The levels of FLG, FLG2, SPRR3v1 and SPRR3v2 transcripts in AD skin were lower than in the control group, while mRNA levels of LELP1, SPRR1Av1, and SPRR1Av2 were significantly higher in the skin of patients with AD compared with the control group. This trend continued even after dividing the results into 3 groups and comparing them: lesional AD skin, non-lesional AD skin and healthy skin from controls, and after excluding the patients with FLG mutations (Fig. 1).
Fig. 1. Level of FLG, FLG2, LELP-1, SPRR1Av1, SPRR1Av2, SPRR1B, SPRR3v1, SPRR3v2 transcripts in skin of atopic dermatitis (AD) and skin of healthy subjects. The level of each mRNA normalized to that of the appropriate reference transcript (ACTB or TBP or G6PD) was determined in lesional (n = 19) and non-lesional (n = 19) skin of patients with AD, and skin of healthy (control) subjects (n = 26).
However, despite the reduced level of FLG transcript in the skin of patients with AD, we observed a positive correlation between SCORAD and the level of FLG transcript in lesional skin. However, the same pattern of changes was observed in FLG2 mRNA in lesional skin, although, in contrast to FLG, the level of FLG2 transcript also increased in non-lesional AD skin as the severity of AD increased (Table I). Moderate correlations of SPRR1Av2 and SPRR3v2 transcript levels were also observed, both in non-lesional AD skin and as the severity of AD increased (Table I). FLG mRNA levels in the skin of patients with AD were higher in patients with elevated IgE, and correlated with the IgE level (Table I). The level of FLG2 mRNA in non-lesional skin, but not in lesional skin, also showed a positive correlation with the IgE level.
Table I. Correlations between transcripts and proteins levels in skin of patients with atopic dermatitis (AD) and clinical variables
The level of SPRR3v2 transcript in the non-lesional AD skin of patients with late-onset AD was higher than the level of that transcript in the non-lesional skin of patients with early-onset AD. A similar trend was observed in lesional skin, but this was not statistically significant (Fig. 2A). Patients with AD and concomitant asthma, when compared with those with AD only, presented lower levels of SPRR1Av1 mRNA in lesional AD skin, as well as in non-lesional AD skin (Fig. 2B).
Fig. 2. Correlation of SPRR3v2 and SPRR1Av1 transcripts levels in skin of patients with atopic dermatitis (AD) with phenotype of disease. (A) SPRR3v2 transcript level was determined in non-lesional and lesional skin of patients with onset of AD at less than 2 years (n = 9) and onset of AD at 2 years or more (n = 10). (B) SPRR1Av1 transcript level was determined in non-lesional and lesional skin of patients with AD with (Yes, n = 10) and without (No, n = 9) concomitant asthma.
The levels of FLG, FLG2, and SPRR3 proteins were significantly lower in AD skin than in the skin of healthy subjects. Moreover, levels of these proteins in lesional focuses of AD skin were distinctly lower than in non-lesional AD skin (Fig. 3). On the other hand, the levels of LELP-1, SPRR1A, and SPRR1B proteins were significantly higher in AD skin compared with the levels of these proteins in the skin of healthy subjects. AD severity inversely correlated with FLG protein level in non-lesional and lesional skin (Table I). Moderate positive correlations between levels of LELP-1, SPRR1A and SPRR1B proteins in non-lesional AD skin and SCORAD were also evident, but these were not statistically significant. The SPRR3 protein level in non-lesional AD skin inversely correlated with pruritus. Some correlations between FLG2, LELP-1, and SPRR1A protein levels in lesional skin and pruritus were also noted, but these relationships did not reach statistical significance (see Table I).
Fig. 3. Level of FLG, FLG2, LELP-1, SPRR1A, SPRR1B, SPRR3 proteins in skin of patients with atopic dermatitis (AD), and skin of healthy subjects. The level of each protein was determined in lesional (n = 6) and non-lesional (n=6) skin of patients with AD, and skin of healthy (control) subjects (n = 14).
Our observations of reduced expression of FLG and FLG2 in the skin of patients with AD are consistent with published data (15–17). They also confirm the results of Pellerin et al. (16), who reported that lowered expression of FLG and FLG2 in AD skin was observed in a subgroup of patients who did not carry FLG mutations. This is in line with another study, which reported that the FLG2 protein was diminished in the presence of inflammatory mediators in AD (18) and might support the outside-inside-outside model of AD pathogenesis. Furthermore, decreased levels of FLG and FLG2 protein in non-lesional AD skin, especially in subjects without FLG mutations, may confirm that the clinically unaffected skin of patients with AD is abnormal and involved in the inflammation process. This phenomenon can also explain the dryness of the skin, especially in the absence of FLG gene mutations (19). Unlike Pellerin et al. (16), who did not find any correlation between FLG protein levels and AD duration or severity, the authors of this study, similar to Batista et al. (17), have noted an inverse correlation of filaggrin expression and disease severity.
It was surprising to find higher mRNA levels of FLG in severe AD compared with moderate AD and a positive correlation between AD severity and elevated FLG mRNA levels accompanied by decreased FLG protein levels, which in turn inversely correlated with AD severity. This observation raises many questions about the molecular mechanisms that, independently of FLG mutations, lead to an elevation of mRNA levels and, finally, result in a lack of protein or a significant reduction in its level in the epidermis. On the other hand, in the light of recent advances in genomics and proteomics, it has become apparent that the mRNA level usually does not predict its protein level (20). Recent studies on thousands of protein/mRNA pairs in several human tissues yielded relatively low Spearman’s correlation coefficients, averaging 0.4 (21). Therefore, it seems that, in the case of FLG in AD skin, the pathway from mRNA to protein warrants further investigation. Moreover, in light of these facts, we propose that both the mRNA and protein levels could be used as independent markers of AD phenotype.
SPRR genes (SPRR1, SPRR2 and SPRR3) encode specialized cornified cell envelope precursor proteins in the stratified epithelia (22, 23). SPRR proteins play a structural role in the cornified envelope as cross-bridging proteins between the loricrins (22, 23). They are recognized as novel cell protective proteins with an important role in wound healing (23), and might play a major role in the adaptation of epithelial barriers to a large variety of external and endogenous stimuli (23, 24). The expression of SPRR proteins increases in response to ultraviolet radiation, ageing, cancer, and wound healing (23). The strong upregulation of SPRR1 in non-lesional AD skin after tape stripping was also observed (25).
All these facts indirectly explain our results, revealing increased expression of SPRR1A in AD skin. Previously, similar findings on SPRR1A expression in AD skin have been reported by other studies (3, 15). However, in contrast to these studies (3, 15), the authors have not observed significant changes in SPRR1B mRNA levels in AD skin; however, they have noted significant elevation of SPRR1B protein level in lesional and non-lesional AD skin. It is worth noting that the level of SPRR1B protein correlated with the SCORAD score. The lack of statistical significance of this relationship may be explained by the small number of cases investigated. Considering the role of SPRR proteins in the epidermis, it seems that up-regulation of SPRR1 might tend to partially compensate for loricrin deficiency (15) in an attempt to repair barrier micro-lesions after scratching or in response to infection. This assumption might be supported by the observation that patients with severe course of AD had higher levels of SPRR1A and SPRR1B mRNA than those with a moderate course of AD. Moreover, levels of SPRR1A protein in nonlesional and lesional skin correlated with AD severity and pruritus, respectively. However, probably owing to a limited number of subjects these correlations did not reach a statistically significant level.
SPRR3 is another cross-bridging protein in the cornified cell envelope (24). The lack of this protein is thought to be responsible for bilayer disorganization (5). Based on the structural differences in the central domain of SPRR proteins, which form a flexible non-ordered structure, it has been postulated that a change in the composition of SPRR proteins could affect the biomechanical properties of CEs (26). This is especially true in the case of the SPRR3 protein, which is more extensively cross-linked than other classes of SPRR proteins (27). Based on this observation, it has been suggested that the SPRR3 protein is an important effector of the biomechanical properties of the cornified envelope (28). The authors found decreased levels of both variants of SPRR3 mRNA in AD skin, accompanied by decreased content of SPRR3 protein. A previous study reported that the SPRR3 mRNA variant carrying an extra 24-bp repeat in the central domain is a risk factor for AD (24). It could be that SNPs cause decreased expression of SPRR3, but this was not confirmed. On the other hand, it is also possible that, similarly to FLG, the expression of SPRR3 in AD skin is diminished by an exogenous stressor or overexpression of Th2 cytokines (11). This warrants further investigation, especially since some of our observations on SPRR3 share similarities with FLG, e.g. decreased mRNA levels of SPRR3 mRNA was associated with early onset of AD, and patients with a severe course of AD presented lower levels of SPRR3 protein in lesional skin than those with moderate AD. Also worth noting is the fact that the SPRR3 protein level inversely correlated with pruritus, which was not observed in earlier studies.
Late cornified envelope-like proline-rich (LELP1) protein is a small, proline-rich protein with an unknown function. According to some authors, LELP1 may be regulated by STAT6, which is responsible for expression of IgE (29). Sharma et al. (29) reported significant association of LELP1 rs7534334 with serum IgE levels in patients with atopic asthma (29). Our previous study showed that CC genotype and C-allele of LELP1 rs7534334 were associated with serum IgE levels, severity of AD and concomitant asthma (30). In the present investigation the authors noted an inverse correlation of LELP1 protein level in non-lesional AD skin with serum IgE level, and a positive correlation with severity of disease and pruritus, but these relationships lacked statistical significance, probably owing to the low number of cases examined.
In summary, our study provides data indicating the correlation of expression levels of CE proteins in the skin of patients with AD with factors that determine the clinical phenotype of this disease. These results point to SPRR3 and LELP1 as potentially new interesting markers of AD. The observed differences and deregulations in protein levels in AD skin might be the result of compensation and protein cooperation, which requires further study.
The authors would like to thank all patients for their participation in the project. The study was supported by grant 2011/03/D/NZ5/00837 from the National Center of Science to MT.
The authors declare no conflicts of interest.


 Click to show fullsize
Click to show fullsize Click to show fullsize
Click to show fullsize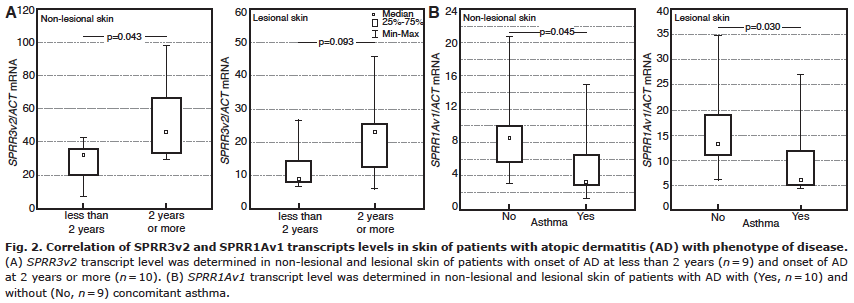 Click to show fullsize
Click to show fullsize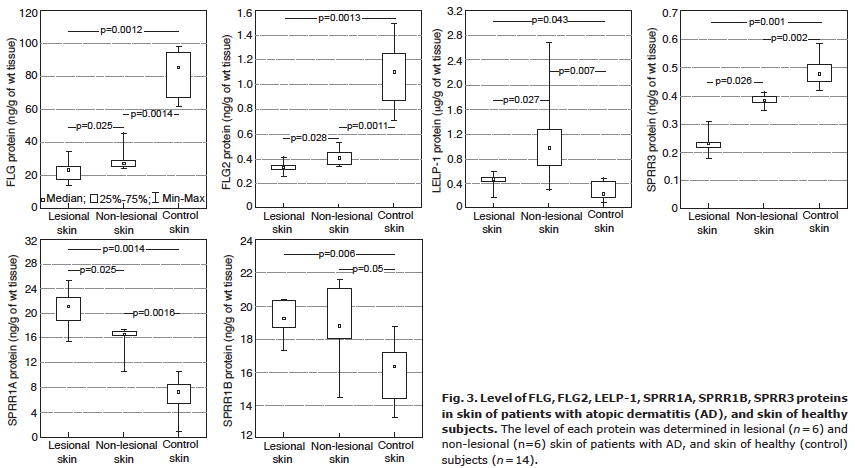 Click to show fullsize
Click to show fullsize