


Departments of 1Dermatology and 3Pathology, University Hospital Zealand, Faculty of Health and Medical Sciences, University of Copenhagen, Roskilde, 2Department of Immunology and Microbiology, Costerton Biofilm Centre, University of Copenhagen, Copenhagen, Denmark, 4Department of Dermatology, Erasmus University Medical Center, Rotterdam, The Netherlands, and 5Department for Clinical Microbiology, Section 9301, Copenhagen University, Copenhagen, Denmark
Hidradenitis suppurativa (HS) is a chronic inflammatory skin disease defined by recurrent nodules, tunnels (sinus tracts) and scarring involving the intertriginous regions. The clinical course of HS is compatible with a biofilm-driven disease, and biofilm has been described in lesional HS skin. We therefore hypothesized that clinically unaffected HS skin would also have an increased presence of biofilm compared with that of healthy controls. We conducted a case-control study, investigating the morphology of the axillary skin microbiota. Peptide nucleic acid – fluore-scence in situ hybridization probes were used in combination with confocal laser scanning microscopy. Significant differences were found in both distribution and quantity of the cutaneous microbiota in clinically non-affected axillary skin of patients with HS compared with healthy controls. Surprisingly, we detected fewer bacteria and less biofilm in patients with HS. The reduced microbiota in patients with HS may play an important role in the early course of the disease.
Key words: hidradenitis suppurativa; microbiota; biofilm; pre-clinical.
Accepted Jul 4, 2015; Epub ahead of print Jul 5, 2016
Acta Derm Venereol 2017; 97: XX–XX.
Corr: Hans Christian Ring, Department of Dermatology, Roskilde Hospital, Køgevej 7-13, DK-4000 Roskilde, Denmark. E-mail: Hrin@regionsjaelland.dk
Hidradenitis suppurativa (HS) is a chronic inflammatory skin disease defined by recurrent nodules, tunnels (sinus tracts) and scarring involving the intertriginous regions. The nodules may expand progressively to abscesses and subsequently rupture, causing suppuration and malodorous discharge (1). The prevalence of HS has been estimated as 1–4% in European populations (2–4), while a mean incidence of 6 per 100,000 person per year has been found in an American study (5). Although the patho-genesis of HS is unknown, the role of the cutaneous commensal bacterial community (microbiota) and its potential relationship to the innate immune system in HS appears to be of increasing interest (6–9). Several studies have hypothesized that a synergistic interaction between the commensal microbiota and aberrant innate immune responses could be a pivotal factor in the pathogenesis of HS (10–12). However, the structure of the bacterial population, as well as the diversity at strain level, is poorly investigated in patients with HS (13–17). Previous studies have furthermore been restricted to lesional skin without inclusion of healthy control groups, and microbiological investigations of clinically normal HS skin of the intertriginous regions are limited.
The clinical course of HS is compatible with a biofilm-driven disease. A biofilm is a microbial-derived, sessile community, characterized by cells attached to a substratum, interface or to each other embedded in a matrix of extracellular polymeric substances (18). Bacterial biofilm infections are clinically intractable infections and notoriously a challenge to eradicate, as biofilm communities exhibit an altered phenotype with respect to growth rate and gene transcription, rendering the bacteria tolerant to antibiotic treatment and the host defence (19). In particular, in chronic wounds, biofilms are thought to delay wound healing and prolong inflammation (20–23).
As recently stated in the European S1 guidelines, the early stage in the pathogenesis of HS is often referred to as an initially sterile process (24). HS is therefore not considered a traditionally infectious disease. However, in lesional skin, biofilm has previously been described and has therefore been suggested to play a role in HS (25, 26). In the 2 studies biofilms were by confocal laser scanning microscopy (CLSM) and fluorescence in situ hybridization (FISH) visualized in sinus tracts and in hair follicles of the lesions. Thus, the pathobiology of HS may elicit formations of bacterial aggregates (biofilms), in particular in anoxic cavities, such as sinus tracts or dilated hair follicles.
If biofilm plays a role in the aetiology of HS, we hypothesized that clinically, unaffected HS skin also would have an increased presence of bacteria and biofilm compared with that of healthy controls. We therefore conducted a case-control study, investigating the axillary skin microbiota comparing patients with HS and healthy controls. Peptide nucleic acid (PNA)-FISH probes were used in combination with CLSM. PNA-FISH probes target specific bacterial ribosomal RNA sequences in the bacterial cells (25, 27). PNA diffuses through the bacterial cell wall due to its uncharged backbone. Importantly, PNA-FISH offers high specificity due to the excellent thermal stability of the RNA-PNA hybrid complexes. Compared with traditional light microscopy and conventional staining methods (e.g. Gram-staining), PNA-FISH in conjunction with CLSM offer high-resolution 3-D images, as well as information about live and dead bacterial cells. Knowledge of the potential variations in the spatial distribution and quantity of the microbiota and biofilm in unaffected HS skin and healthy controls may offer novel insights into the early pathogenesis of HS.
This study was accepted by the ethics committee of Region Zealand (project number SJ-420) and the data protection agency (REG-105-2014) in Denmark. The study was conducted at Roskilde Hospital, Department of Dermatology from October 2014 to August 2015. Written informed consent was obtained from all study participants.
An exploratory case-control study comparing individuals with a diagnosis of HS vs. healthy controls (n = 24, respectively) was performed.
All cases were consecutive HS patients the from Department of Dermatology, Roskilde Hospital. Healthy controls were recruited at the University of Copenhagen and among the staff at the Department of Dermatology, Roskilde Hospital.
Inclusion criteria. All cases had a verified diagnosis of HS (ICD-10 code: L732) at the Department of Dermatology, Roskilde Hospital. All were aged over 18 years and non-pregnant. All HS patients reported intermittent activity of inflamed nodules in both axilla. Thus, all included patients had a history of previously active lesions at the investigated site. However, all biopsies contained only clinically normal-appearing skin (without scarring, inflammation or swelling) with clinically visible hair follicles.
Inclusion criteria of healthy controls. No known skin disease. No visible skin disease in axillae, in particular no folliculitis, nodules, tunnels (sinus tracts), abscesses or scars compatible with current or previous HS lesions.
Exclusion criteria of cases and controls. Treatment with antibiotics within one month prior to the study (systemic or topical therapy) or current flares (inflamed lesions, cases).
Other medications. Seven of the included cases administered themselves with topical Resorcinol in the groin. Healthy controls were not on any other type of medications at the time of investigation. We did not obtain specific information on oral contraceptives among patients with HS or healthy controls.
Prior to injection of anaesthetics, the skin was cleansed with ethanol swabs (FastCare®, 70% Isopropyl Alcohol, Zhejian, China) to reduce the number of surface microorganisms.
Axillary skin was anesthetized using a 2% solution of lidocaine and adrenaline (no preservatives). A 4-mm punch biopsy was obtained in the axilla of patients with HS and healthy controls. The biopsy was stored in formalin and subsequently paraffin embedded according to the standard operating procedure of the pathology department, Roskilde Hospital. In order to identify the pilosebaceous units each paraffin block was sectioned at 5+ levels, each level consisting of 7 slides (4 µm thick), and processed until a representative pilosebaceous unit was identified. The slide in the middle group was stained with haematoxylin and eosin (HE) and examined by standard light microscopy for the presence of representative areas of pilosebaceous units and subsequent identification of bacteria and grading of inflammation. The pathologist who performed the HE investigations was blinded.
Paraffin-embedded tissues were deparaffinized using a standardized method (27): (i) xylene (2 × 5 min), (ii) 99% ethanol (2 × 3 min.), (iii) 96% ethanol (2×3 min.), (iv) Milli-Q water (3 × 3 min), incubated in 90 min at 55°C with PNA-FISH-TexasRed (red) conjugated (a specific universal bacterial (UniBac) 16S ribosomal RNA) probe (AdvanDx, Woburn, MA, USA). Additional staining by a specific coagulase-negative staphylococci (CNS) PNA-FISH probe was performed on slides categorized as positive by the use of the specific Unibac probe. The slides were incubated in wash buffer (AdvanDx, Woburn, MA, USA) for 30 min at 55°C and subsequently counter-stained with 4’,6-diamidino-2-phenylindole (DAPI) (Life Technologies, Oregon, USA) (3 µM) for 15 min at RT, rinsed in Milli-Q water, added mounting media (Prolong Gold, Life technologies, Oregon, USA) and subsequently mounted on a coverslide and sealed with clear nail polish.
Two investigators preformed the CLSM analysis unblinded. The PNA-FISH/DAPI stained slides were examined by CLSM (Axio Imager.Z2, LSM710 CLSM; Zeiss, Oberkochen, Germany) and the accompanying 3D reconstruction software (Zen 2010, version 6.0; Zeiss, Oberkochen, Germany). Fluorescence images were recorded at 405, 488 and 594 nm excitation wavelength and an emission wavelength at 410–483 nm (blue), 492–589 nm (green) and 599–690 nm (red), respectively. The lasers were used in combination with the corresponding beam-splitters MBS 488/594 and MBs_InVin 405 (Zeiss, Oberkochen, Germany). The objectives used were 40×/1.3 (numerical aperture) Plan-Neoflour and 63×/1.4 plan-apochromat oil objectives (Zeiss, Oberkochen, Germany). As the remaining non-stained tissue appeared weak green by autofluorescence, the images were taken in 1 track and thus intensified the green signal by the use of all 3 lasers. The images had a resolution of 1,024×1,024 pixels and a colour depth of 16 bits. Each pixel was scanned twice and subsequently deconvoluted into Tiff-files (Imaris 8 ×64 version 8.1.2, Bitplane, Zürich, Switzerland).
In addition to standard light microscopy, CLSM were performed on a minimum of 3 slides; 1 or 2 on each side of the HE slide. With regard to the classification of positive vs. negative samples, a definition of bacterial aggregates was applied. In a recently published study, bacterial aggregates exceeding 5 µm in diameter were categorized as biofilm (28). The given classification for quantitative assessment of the bacterial aggregates was subsequently defined by the following: 0 = no bacteria detected or single scattered cells < 10, 1 = 5–10 µm aggregates or single scattered cells > 10, 2 = 10–50 µm aggregates, 3 > 50 µm aggregates. For each of the positive samples, the bacterial morphology and the location of the findings was noted. The entire PNA-FISH slide was reviewed. Overall, the PNA-FISH CLSM results reflect that specific slide from each participant that yielded the highest positive score in aggregate size.
Due to staining with the PNA-FISH probe and DAPI, active bacteria were visualized as bright-red or pink in a combined microscope filter, whereas more dormant bacteria appeared purple or blue due to the lower level of rRNA present in the cells (29).
In addition, the fluorescence-stained slides were scanned using a digital slide scanner (Axio scan. Z.1, Zeiss, Oberkochen, Germany) with a 20×/0.95 objective (Zeiss, Oberkochen, Germany) providing an optimized overview of the slides.
The frequent distribution of bacterial aggregate quantification between the 2 groups of participants was analysed by Fischer’s exact test. In addition, a χ2 test was performed on the location of bacterial aggregates. The criteria for statistical significance was p < 0.05. The analyses were performed in GraphPad Prism 5.02 software (GraphPad Software Inc., California, USA).
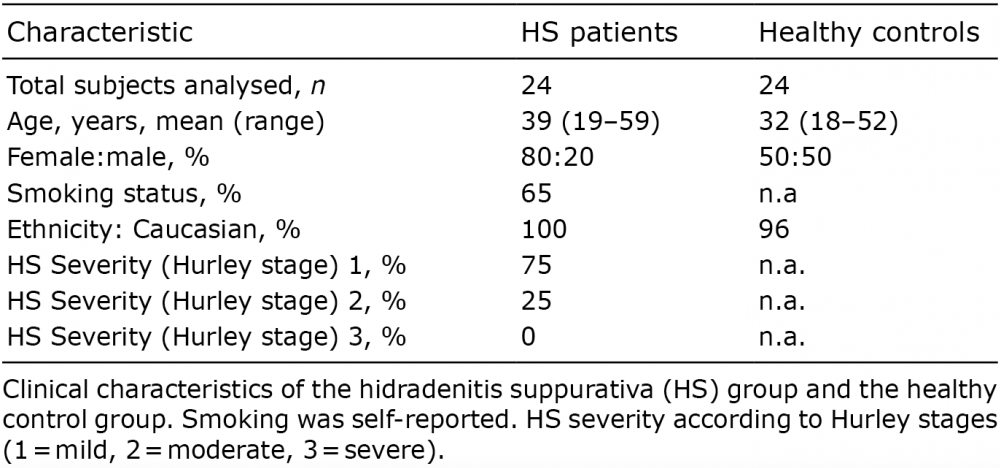
Our study comprised 24 HS cases and 24 healthy controls. The background factors and characteristics of the HS group and controls showed that a majority of the patients with HS were predominately younger, female and smokers (Table I). For each participant 4 histological slides (3 CLSM samples (Universal PNA-FISH) and 1 HE sample) were examined. With additional CNS PNA-FISH staining of Universal PNA-FISH-positive samples (n = 25 positive), the overall investigation yielded, in total, investigations of 217 slides.
Table I. Background factors and characteristics
Overall, PNA-FISH and CLSM examinations of pre-clinical HS skin showed an absence of bacterial aggregates at the stratum corneum (SC) and in hair follicles compared with healthy controls. As shown in Table II, only 12% of the HS samples were categorized as positive for small aggregates or single scattered cells. The 12% of the HS patients who had bacterial aggregates were 2 females and one male. At the time of investigation, none of the 3 patients was on any medications. Thus, the majority of the HS samples showed no bacterial aggregates (Figs 1 and 2). In comparison, PNA-FISH and CLSM examinations demonstrated morphologically significant presence of bacterial aggregates (biofilm formations) in 92% of the healthy controls. The majority of the aggregates were found in hair follicles (64%) or in the SC (36%) (Table II and Figs 3 and 4).
Table II. Bacterial quantification, distribution, morphology and histopathology in patients with hidradenitis suppurativa (HS) vs. healthy controls
Fig. 1. Distribution and quantification of bacterial aggregates in pre-clinical hidradenitis suppurativa (HS) patients vs. healthy controls. (a) In healthy controls, bacterial aggregates were found in the majority of the samples, primarily situated in the hair follicle (HF) or at the stratum corneum (SC). In the pre-clinical HS a high occurrence of negative samples were found. Only 3 samples were found positive for small bacterial aggregates. (b) Quantification of bacterial aggregates in skin biopsies shown in a dot plot with median of 0 (zero) and of 2 for the HS patients and controls, respectively. Each dot represents the quantification of one sample. ***p < 0.001 for controls vs. patients.
Fig. 2. Slide scan and confocal laser scanning microscopy images of negative hidradenitis suppurativa case. (a) Slide scan of hair follicle (HF) (200× magnification). The tissue appears green due to auto-fluorescence, while the human nuclei are stained blue by 4’,6-diamidino-2-phenylindole (DAPI). A strand of hair is visible in the inferior segment of the HF (white arrow). Keratinous material appears within the lumen of the HF. (b) Magnification of the same HF showing the infundibulum and the isthmus. No bacteria are detected (they would have appeared pink/blue). Erythrocytes appear yellow (auto-fluorescence) at the infundibulum (white arrows). All slides were stained with DAPI (blue) and peptide nucleic acid – fluorescence in situ hybridization UniBac with TexasRed (red).
Fig. 3. Confocal laser scanning microscopy (CLSM) images of control sample. (a) Stratum corneum with several bacterial aggregates (biofilm, pink colour) marked by white arrows. In (b) and, (c) magnification of the area, CLSM images visualize the morphology of the bacterial aggregates (white arrows).
Fig. 4. Slide scan, haematoxylin and eosin (HE) and confocal laser scanning microscopy (CLSM) images of a healthy control. (a) Slide scan of hair follicle (HF) (200× magnification). (b) Corresponding HE staining, bacterial aggregates (biofilm) appear dark purple in the centre of the HF. In (c) and (d) magnification of the area. The CLSM images depict the massive presence of biofilm within the lumen of the HF (arrows).
The morphology of the bacterial aggregates were predominantly classified as cocci (92%) whereas rods constituted only a minor percentage (8%) (Table II). Of the 92% categorized as positive by the use of the Universal PNA-FISH probe, only 35% of the samples were positive for CNS bacteria (Table II).
Quantification of the bacterial aggregates showed mainly measures of 10–50 µm (diameter) (Table II and Fig. 4). In general, there was a tendency for more active bacterial cells to be present in SC, while bacteria present in the hair follicles were a mixture of either active and dormant or pure dormant cells (29).
Statistical analysis on the qualitative validation of bacterial aggregates yielded a significantly decreased occurrence in HS cases compared with healthy controls (p < 0.0001). In addition, a significant difference was found in the distribution of aggregates in relation to location (p < 0.0001).
Histopathological investigations of the biopsies of HS cases and controls were in agreement with the CLSM findings, showing good morphological correlation between biofilm aggregates found in HE slides and the corresponding CLSM images (Fig. 4). According to clinical routine practice, depending on the presence of lymphocytes, the inflammation was graded on a scale ranging from 0 to 3 (none, mild, moderate or severe).
Although all biopsies were obtained from clinically unaffected skin, a significant proportion of HS samples (50%) showed signs of low-grade inflammation (mild presence of lymphocytes). In comparison, only 16% in the control group presented low-grade inflammation. A detailed examination of the inflammation among the HS cases revealed widely scattered lymphocytic infiltrates, mast cells, dilated hair follicles and keratin plugging of the infundibulum. Occasionally, plasma-cells, neutrophils and eosinophils granulocytes were seen in both groups.
Our study demonstrated significant differences in the distribution and quantity of the cutaneous microbiota as well as biofilm formations in clinically non-affected axillary skin of patients with HS compared with healthy controls (Fig. 4), with fewer bacteria and less biofilm found in HS. The observations are in sharp contrast with our initial hypothesis of an increased presence of biofilm in clinically unaffected HS skin. Nevertheless, these results may give rise to new pathogenic concepts for HS. The homogeneity observed in the 2 groups is statistically significant, and it is therefore suggested that the results may be extrapolated to HS populations and healthy individuals on a larger scale.
PNA-FISH and CLSM are often used as the gold standard when spatial insights on the distribution of bacterial aggregates are required (27, 28, 30–32). Although, PNA-FISH and CLSM does not depict the extracellular matrix of the bacterial aggregates, the applicability of the technique has been demonstrated in various skin diseases (25, 31, 33–35). Previous studies have suggested a possible pathogenic role for biofilm in the development of HS (25, 26). A histological retrospective study of biopsies from lesional skin in 27 patients with HS was performed using CLSM and FISH (25). Biofilm formations were seen in one-fifth (20%) of the samples and were primarily situated in hair follicles and sinus tracts. However, taking our recent findings into account the presence of biofilm found in lesional HS skin may thus primarily reflect secondary colonization of biofilm-producing bacteria, most likely due to rupture of the innate skin barrier. This also appears to be in agreement with the bacterial findings from lesional HS skin, often revealing a wide spectrum of commensal bacteria (biofilm producing), such as CNS bacteria (8). This study also verified the presence of CNS in the axilla of healthy controls (35%) (Table II).
Apart from the findings in the HS group, our study also importantly demonstrated the normal spatial distribution as well as quantification of bacterial aggregates in healthy
axillary skin using CLSM and PNA-FISH. To the best of the author’s knowledge, the healthy microbiota in axillary skin has not previously been visualized by the use of CLSM and may therefore be used in other microbiome studies for comparison.
Considering the fact that the intertriginous regions of the human skin and, in particular, the axilla, offers highly favourable conditions for bacteria to proliferate (36), our observation of a reduced presence of bacterial aggregates in axillary HS skin appears even more interesting. Remarkably, alterations in the composition of the skin microbiota (dysbiosis) has been shown in the context of other skin inflammatory disorders, such as acne vulgaris, psoriasis and atopic dermatitis (37). Thus, it may be speculated whether an altered shift in the resident microbiota may be implicated in the patho-genesis of HS.
Although bacteria are often referred to as catalysers of inflammation, recent studies have demonstrated that symbiotic factors produced by the cutaneous and intestinal commensal microbiota beneficially modulate the host immune systems, i.e. by regulating inflammatory responses (38–43). Thus, substantial evidence supports the hypothesis that the normal skin microbiome acts as an antimicrobial shield and contributes to normal defence at the epidermal interface. It may therefore be speculated whether the altered microbiota (lack of biofilms) in pre-clinical HS leave the patients more susceptible to invading pathogens. Alternatively, the local dysbiosis may trigger immune-mediated inflammation.
The major advantages of our study are the design (case-control study) and the extensive investigation of the skin samples. Furthermore, there is minimal misclassification bias, as an experienced dermatologist diagnosed the HS cases. Although, routine ethanol swabs were applied prior to sampling (biopsy) of the skin, remarkable biofilm formations (containing both active and dormant cells) were seen at the SC (Fig. 3), primarily consisting of cocci (92%). Furthermore, the healthy control group also exhibited massive biofilm formations within the hair follicles (Fig. 4).
Some limitations may have influenced the results. Although deodorants and other antiperspirants may influence species diversity (44), we did not obtain information on personal hygiene or intervals between showering and sample collection. Furthermore, the HE and PNA-FISH findings were not validated with culture data. The HS participants were not sex-matched with the healthy controls and potential HS subtypes were not stated. The information on use of antibiotics was self-reported, yielding possible recall bias. Moreover, we sampled only once and did not check the development of biofilm over time. We have chosen a minimum of one month after the last antibiotic treatment as a cut-off, which is in agreement with a previously published guideline on optimized sampling methods in HS (7). In addition, we did not visualize the extracellular matrix of the bacterial aggregates.
We detected fewer bacteria and less biofilm, indicating a relative lack of skin microbiota in clinically unaffected axillary HS skin. The reduced microbiota in patients with HS may play an important role in the early course of the disease. Moreover, the high level of presence of cocci-biofilm (92%) seen among the healthy controls highlights the potential biological importance of cocci species. Future follow-up studies may determine the development of the microbiota over time. Furthermore, general investigations of the complex cross-talk between the resident microbiota and epithelial cytokines in the skin are needed to clarify the mechanisms that modulate skin immune homeostasis and subsequent inflammation.
The authors would like to thank The Lundbeck Foundation for financial support of Professor Thomas Bjarnsholt.
The authors declare no conflicts of interest.


 Click to show fullsize
Click to show fullsize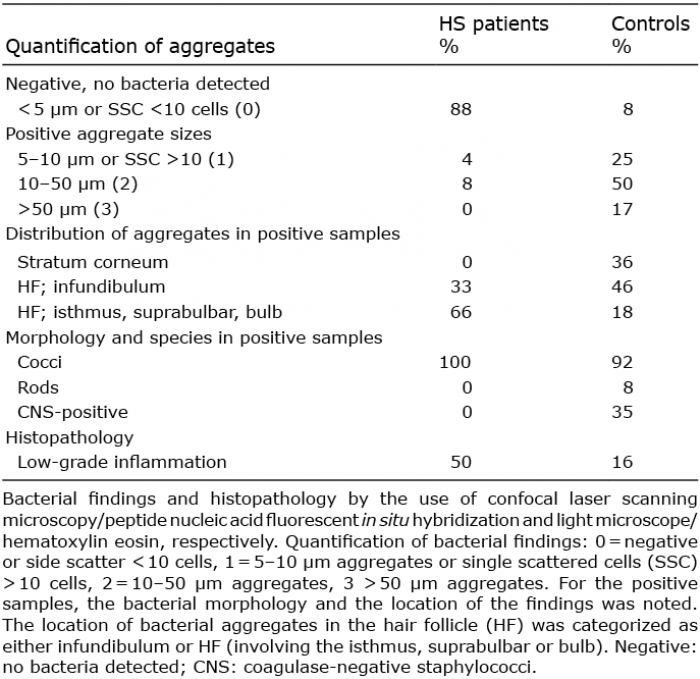 Click to show fullsize
Click to show fullsize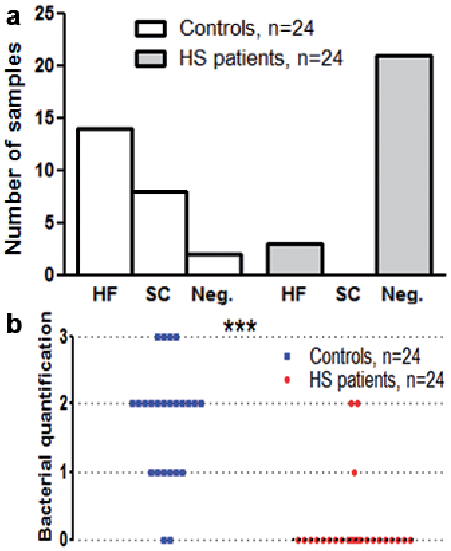 Click to show fullsize
Click to show fullsize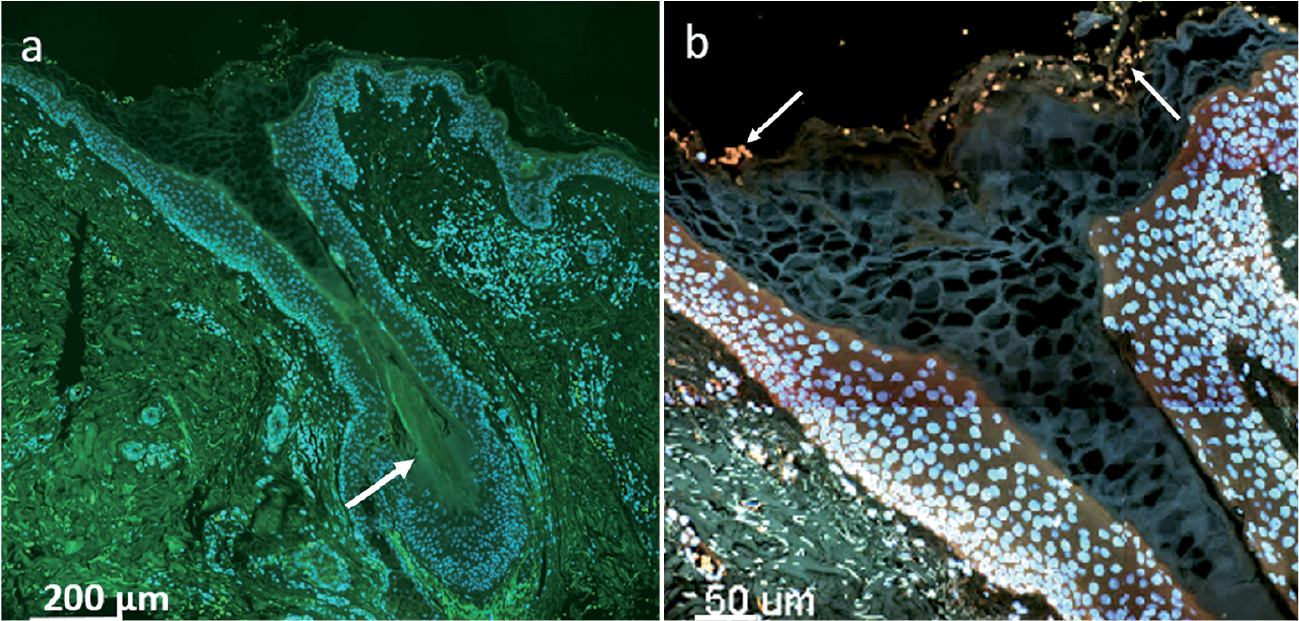 Click to show fullsize
Click to show fullsize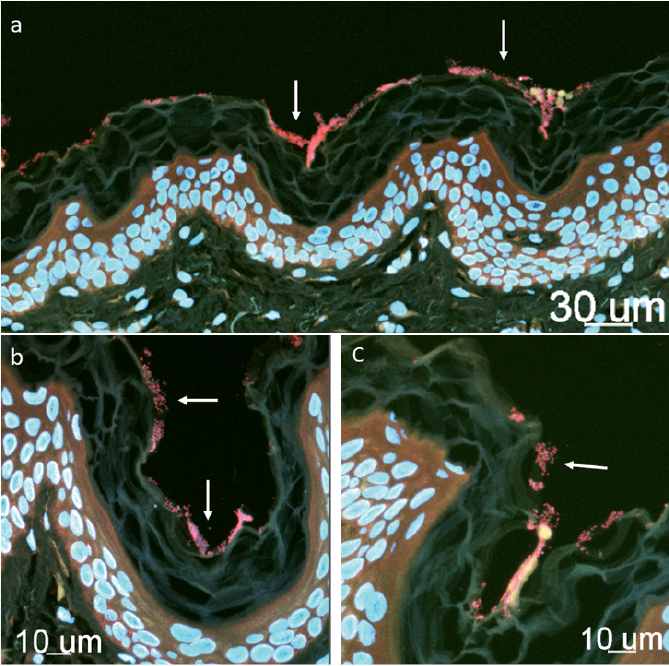 Click to show fullsize
Click to show fullsize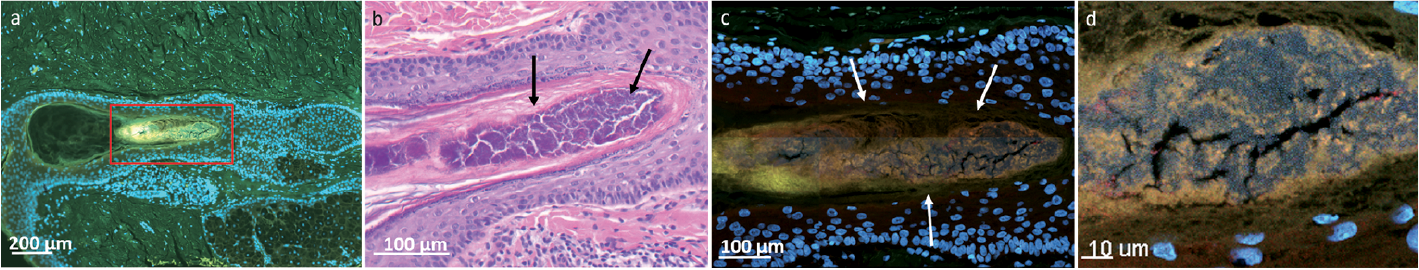 Click to show fullsize
Click to show fullsize