


1Departments of Dermatology, Venereology, Allergology and Immunology, Dessau Medical Center, Dessau, Germany, 2ASI Corporation, Bridgewater, USA, and 3Delhi Dermatology Group, Kubba Clinic, New Delhi, Delhi, India
*Preliminary results of this work were presented as posters at the Society for Investigative Dermatology 2005, Arbeitsgemeinschaft Dermatologische Forschung 2012 and the European Society for Dermatological Research 2012 meetings, and as a lecture at the Entretiens d’Avène 2014.
Calcium and 1,25 dihydroxyvitamin D3 (1,25(OH)2D3) are promoters of epithelial cell functions; however their effects on sebaceous glands are unknown. In this study, morphology, ultrastructure, cell numbers, lipid synthesis and apoptosis of SZ95 sebocytes were assessed in vitro under different concentrations of extracellular calcium with or without 1,25(OH)2D3. Moreover, serum calcium and 1,25(OH)2D3 levels were assessed in acne and non-acne patients (controls). Under conditions of low extracellular calcium, lipogenesis and cell detachment were observed. Increasing extracellular calcium enhanced sebocyte numbers, induced epithelial mor-phology and reduced lipogenesis. Moreover, a reduction in extracellular calcium reduced E-cadherin and enhanced caspase 3/7 activity (apoptosis), whereas calcium chelation by EGTA (ethylene glycol-bis(β-aminoethyl ether)-N,N,N’,N’-tetraacetic acid) resulted in enhanced lipogenesis. 1,25(OH)2D3 decreased sebaceous lipogenesis, but also induced signs of autophagy. In the clinical study, patients and controls exhibited normal serum calcium levels, whereas younger acne patients presented higher levels than older patients and controls. In addition, younger acne patients presented lower 1,25(OH)2D3 levels than did older ones. In conclusion, extracellular calcium and 1,25(OH)2D3 regulate sebocyte morphology, increase cell numbers, decrease sebaceous lipogenesis and induce cell autophagy in vitro. The increased ionized calcium and the reduced 1,25(OH)2D3 levels detected in the serum of younger patients with acne may contribute respectively to increased sebaceous gland volume and enhanced lipogenesis.
Key words: calcium; vitamin D; dihydroxyvitamin D3; sebocytes; differentiation; proliferation; lipids; autophagy; acne.
Accepted Aug 29, 2016; Epub ahead of print Aug 30, 2016
Acta Derm Venereol 2017; 97: XX–XX.
Corr: Christos C. Zouboulis, Departments of Dermatology, Venereo-logy, Allergology and Immunology, Dessau Medical Center, Auenweg 38, DE-06847 Dessau, Germany. E-mail: christos.zouboulis@klinikum-dessau.de
1α,25-dihydroxyvitamin D3 (1,25(OH)2D3) was studied in the treatment of acne by Maynard as early as 1938 (1), followed by Cerri (2), Carli (3) and Garnier (4). However, to our knowledge, no previous clinical studies have been performed on serum levels of 1,25(OH)2D3 in patients with acne, and the use of vitamin supplementation in the absence of obvious deficiency has been logically criticized (5). In a current systematic review of off-label uses of topical 1,25(OH)2D3 in dermatology, seborrhoea and acne were not among the disorders evaluated (6). Moreover, there are no references on serum levels of calcium (Ca2+) in acne, despite the fact that, in several studies, treatment of acne with isotretinoin has been shown to modify circulating 1,25(OH)2D3 and Ca2+ levels (7–10).
Calcium ions play a major role in the regulation of keratinocyte growth and differentiation. Normal keratinocytes cultured in low extracellular Ca2+ (ecCa2+) do not differentiate, whereas they proliferate rapidly. On the other hand, high ecCa2+ increases intracellular Ca2+ (icCa2+) and, together with 1,25(OH)2D3, regulates a sequential turning on and off of the genes possessing response elements, as well as activating the enzymes that induce keratinocyte differentiation and trigger apoptosis (11). The modulation of icCa2+ in vitro, also through 1,25(OH)2D3 (12), has been a useful tool to illuminate keratinocyte biology (12) and various related skin diseases, such as Darier’s disease and Hailey-Hailey disease, where mutations in Ca2+ ATPases (SERCA2, SPCA1) have been detected (11).
Beyond keratinocytes, Ca2+ is increasingly being recognized as a central signal transmitter in other cells, including sebocytes (13). Sebaceous glands are specialized skin appendages committed to differentiate, synthesize and secrete lipids, which, together with cell debris, form the sebum. Sebocytes are grouped into glandular acini, which are composed of an outermost basal proliferating layer and suprabasal differentiating layers that gradually accumulate lipids and finally rupture (holocrine secretion) (14, 15), found to undergo apoptosis prior to cell bursting and death (16). Interestingly, the marked stimulatory effect of arachidonic acid on sebaceous lipogenesis (17) was almost completely abrogated by lowering ecCa2+ levels to 0.25 mM (18). In addition, α-melanocyte-stimulating hormone increases lipogenesis in SZ95 sebocytes (19, 20), and regulates icCa2+ levels, in parallel (19). Moreover, icCa2+ chelation induced apoptosis in hamster sebocytes (21). Al-though keratinocytes and sebocytes are both epithelial cells, they are antipodes regarding their mode of differentiation, as shown in our previous studies (22, 23). Progenitor epithelial cell has to “decide” during their differentiation between a keratinocyte or a sebocyte lineage pattern (24).
SZ95 sebocytes express the vitamin D receptor (VDR) at mRNA and protein levels and possess all enzymes required for the synthesis and metabolism of 1,25(OH)2D3, the biologically active metabolite of vitamin D (25). An anti-inflammatory effect of 1,25(OH)2D3 on human sebocytes has been shown, as 1,25(OH)2D3 reduced interleukin (IL)-6 and IL-8 as well as metalloproteinase-9 expression and secretion (25, 26) at ecCa2+ levels of 0.4 mM (21). Since neither the exact “Ca2+ switch” concentration in human sebocytes nor the ecCa2+ levels/1,25(OH)2D3 interaction on sebocyte differentiation, lipogenesis, and apoptosis are known, it was the aim of our in vitro study to elucidate these events. In addition, a clinical study was conducted in order to evaluate Ca2+ and 1,25(OH)2D3 levels in patients with acne and to detect the clinical relevance of the in vitro results.
Immortalized human SZ95 sebocytes shown to maintain the major characteristics of normal sebocyte differentiation (27) were cultured in human sebocyte growth medium (Sebomed®; Biochrom, Berlin, Germany) supplemented with 10% foetal calf serum (FCS), 5 ng/ml human recombinant epidermal growth factor, 50 µg/ml gentamicin and 1 mM CaCl2 (all from Biochrom) until reaching 70% confluence. The cells were then detached with trypsin/ethylendiamine tetraacetic acid 0.05%/0.02% and transferred to 96-well plates at a density of 8,000 cells/well (assessment of cell number), 10,000 and 20,000 cells/well (detection of apoptosis) and 20,000 cells/well (evaluation of lipid synthesis). Cells were maintained in supplemented human sebocyte growth medium overnight at 37ºC and 5% CO2 and after washing twice with phosphate-buffered saline without Ca2+ and Mg2+ (PBS; Biochrom), they were switched to serum-free sebocyte growth medium (SFM) containing 5 ng/ml hEGF, 50 µg/ml gentamicin, 10–6 M linoleic acid, 10 µg/ml insulin, 1 mg/ml bovine serum albumin and 10–6 M retinol and supplemented with Ca2+ at different concentrations according to the requirements of the experiment.
Since SZ95 sebocytes have been maintained in Ca2+-containing culture medium over several subcultures, the cells were handled for 24 h under the culture conditions described above, with the addition of the potent extracellular calcium chelating agent ethylenglycol-bis(aminoethylether)-N,N,N′,N′-tetraacetic acid (EGTA; Sigma-Aldrich, Deisenhofen, Germany) at concentrations of 0.1–0.6 mM in 1M NaOH.
SZ95 sebocytes were cultured as described above and SFM was supplemented with Ca2+ at concentrations of 0.05, 0.2, 0.4, 0.8, 1.0 and 1.4 mM. Changes in cell morphology were documented 24 h after seeding (day 0) and every 3 days thereafter for 12 days with an Olympus CK 40 phase contrast microscope (Hamburg, Germany).
For transmission electron microscopy SZ95 sebocytes were grown in culture dishes with and without EGTA for 24 and 72 h. Cells were then fixed in modified Karnovsky’s fixative solution for 1 h at room temperature, scraped off, washed with 0.1 M cacodylate buffer and pelletted in microcentrifuge tubes. The pellets were embedded first in agar and the resulting samples were post-fixed in 1% OsO4, dehydrated routinely through a series of graded ethanol and embedded in a low viscosity epon-epoxy mixture. Ultrathin sections cut using a diamond knife were double stained with uranyl acetate and lead citrate and observed under an electron microscope at 60 KV.
To study the effect of low and high ecCa2+ conditions, SZ95 sebocytes were left to attach for 24 h, then washed twice with PBS and half of the cultures were maintained for 48 h in SFM at 0.05 mM Ca2+ (low ecCa2+ conditions) and half in SFM supplemented with 1.4 mM Ca2+ (high ecCa2+ conditions). Finally, the cells were washed twice with PBS and switched to SFM supplemented with Ca2+ at concentrations of 0.05, 0.2, 0.4, 0.8, 1.0, or 1.4 mM. After 24 h, the fluorescein diacetate (FDA) assay for detecting cell numbers and the Nile red (NR) fluore-scence assay for identifying intracellular lipids were performed as described previously (27, 28). Three complete independent experiments, each in sixplicate wells, were performed.
SZ95 sebocytes were left to attach for 72 h, washed twice with PBS and switched to SFM supplemented with Ca2+ at concentrations of 0.05, 0.2, 0.4, 0.8, and 1.4 mM for 24 h at cell confluence. Caspase-3/7 activity, a marker of cell apoptosis, was detected using a commercial assay (Apo-ONE Homogenous Caspase-3/7 Assay; Promega, Madison, WI, USA) according to the manufacturer’s instructions. The released fluorescence was read on a Molecular Devices SPECTRA Max Gemini spectrofluorometer with 499 nm excitation and 521 nm emission filters. The experiment was performed twice.
For protein extraction, SZ95 sebocytes were harvested after incubation with Ca2+ (24 h: 0.05, 0.2, 0.4, 0.8, 1, and 1.4 mM; 72 h: 0.05, and 1.4 mM) and lysed in radioimmunoprecipitation assay buffer (RIPA) buffer supplemented with complete protease inhibitor and phosphatase inhibitor cocktail (Roche, Penzberg, Germany). Extracts were homogenized and centrifuged at 12,000 × g for 10 min. Sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) and Western blot analysis of total proteins were performed as described previously (29). Immunoblotting was performed with rabbit monoclonal antibodies against E-cadherin (1:1000; Cell Signaling, Danvers, MA, USA; cat. no. 3195) and β-actin as 1: 500; control (Cell Signaling; cat. no. 4670). After incubation with a peroxidase-labelled goat anti-rabbit antibody (1:5000; Dako, Hamburg, Germany), antigen–antibody complexes were detected by enhanced chemiluminescence (ECL) Western blotting detection reagents (Peqlab, Erlangen, Germany). One to 30 min exposures were analysed with the Fusion FX7 imaging system (Peqlab).
In order to detect E-cadherin activity, SZ95 sebocytes were stained employing immunocytochemistry. Cells were first fixed in PBS containing 4% paraformaldehyde (Science Services, Munich, Germany) for 20 min at room temperature, subsequently washed twice with PBS and finally blocked with 10% FCS and 0.1% Triton X-100 (Sigma-Aldrich) in PBS. Staining with the E-cadherin monoclonal antibody was followed by an Alexa-488-conjugated secondary antibody (1: 300, Invitrogen, Karlsruhe, Germany; cat. no. A11001). Nuclei were counter-stained with 200 ng/ml 4′,6-diamidin-2-phenylindol (DAPI; Invitrogen) and visualized using fluorescence microscopy.
SZ95 sebocytes were cultured as described above and SFM was supplemented with Ca2+ at concentrations of 0.05, 0.2, 0.4 and 1.4 mM. In each setting, the medium was supplemented with 1,25(OH)2D3 at concentration of 10–9 and 10–7 M. After 48 h and 72 h FDA and NR assays were performed for assessment of the sebocyte numbers and intracellular lipids, as described above.
India was chosen for the location of the clinical study, since there is no major difference in sunshine hours between summer and winter. The serum of 104 Indian patients with acne and 112 patients with other dermatological diseases with no history of acne, serving as controls (Table SI), was drawn after providing a consent for examination of Ca2+ and 1,25(OH)2D3 levels by commercial assays. Serum was drawn in 79 untreated patients, and in 25 patients at the start of treatment. The clinical severity of acne was assessed as mild, moderate or severe.
The in vitro experiments were performed 3 times, except where otherwise reported. The statistical significance of the in vitro data was evaluated with the paired 2-tailed Student’s t-test. All in vitro data are presented as mean values ± standard deviation (SD). In vivo data were evaluated by non-parametric tests and are presented as median and 95% confidence intervals (95% CI). Statistical evaluation was performed by the statistical product and service solutions software (SPSS, Chicago, IL, USA) for Windows (version 18). A p-value of < 0.05 was considered significant.
SZ95 sebocytes maintained under low ecCa2+ levels (0.05 mM) exhibited a rounded to polygonal shape, mostly remained as single cells or formed few loose colonies that poorly expanded and tended to detach from culture plates and to float in the culture medium. On the other hand, SZ95 sebocytes maintained under high ecCa2+ levels (1.4 mM) were polygonal in shape and formed colonies. Despite the absence of serum, SZ95 sebocytes under high ecCa2+ levels reached confluence on day 6 of incubation (Fig. 1).
Fig. 1. Morphology of SZ95 sebocytes at days 1 (a, b), 3 (c, d) and 6 (e, f) under high (a, c, e; 1.4 mM) and low (b, d, f; 0.05 mM) ecCa2+ levels, respectively. Human SZ95 sebocytes incubated at low ecCa2+ levels show loose cell-to-cell contact, stay as single cells or form 2–3 small cell colonies in the culture plates and most of them detach and are floating in the culture medium. SZ95 sebocytes maintained at high ecCa2+ levels grow rapidly in monolayer colonies and reach confluence at day 6 (original magnification × 100).
Electron micrographs of SZ95 sebocytes maintained in SFM at low ecCa2+ levels (0.05 mM) showed numerous mitochondria, highly developed Golgi complex and several small to large lipid droplets consistent with active cell metabolism and lipogenesis (Fig. 2 a,b). In contrast, SZ95 sebocytes maintained at ecCa2+ concentrations ≥ 0.4 mM showed a predominantly undifferentiated morphology and no lipogenesis could be detected (Fig. 2 c,d).
Fig. 2. Ultrastructure of SZ95 sebocytes depending on ecCa2+ levels. Electron micrographs of sebocytes maintained at (a, b) low ecCa2+ levels (0.05 mM) show numerous mitochondria (M), highly developed Golgi complex (G) and several small to large lipid droplets (LD) consistent with active cell metabolism and lipogenesis. ER: endoplasmic reticulum; (c) high ecCa2+ levels (0.4 mM) show a predominantly undifferentiated morphology, whereas lipid droplets are barely detected (d). N: nucleus. (e) SZ95 sebocytes maintained at high ecCa2+ levels (1.4 mM) with the calcium chelator EGTA (ethylene glycol-bis(β-aminoethyl ether)-N,N,N’,N’-tetraacetic acid) for 48 h show a highly developed Golgi complex (G) and increasing small lipid droplets (SLD) consistent with an upregulation of lipid synthesis. (f) SZ95 sebocytes maintained at high ecCa2+ levels (1.4 mM Ca2+) and 1,25(OH)2D3 (10–7 M) showing autophagic vesicles (AV), microvilli (MV) and lipid droplets (LD). (g) SZ95 sebocytes maintained at low ecCa2+ levels (0.05 mM Ca2+) and 1,25(OH)2D3 (10–7 M) show late stage autophagic vesicles (AV). The large vacuoles are extracellular spaces as indicated by the microvilli (MV) often sectioned obliquely.
The increase in ecCa2+ levels was associated with a significant increase in SZ95 sebocyte numbers in a dose-dependent manner during the short experimental period (24 h). Interestingly, a significant increase in cell numbers was observed at all ecCa2+ levels examined, namely 0.2 (p < 0.01), 0.4 (p < 0.01), 0.8 (p < 0.001), 1.0 (p < 0.001) and 1.4 mM (p < 0.001) in comparison with SZ95 sebocytes maintained at 0.05 mM ecCa2+ (Fig. 3).
Fig. 3. Effect of ecCa2+ levels on SZ95 cell numbers at 24 h. The results presented, performed in sixplicate, are from 1 out of 2 representative experiments performed and are calculated as the absolute fluorescence units (AFU) of each experiment. **p < 0.01; ***p < 0.001.
In contrast, the decrease in ecCa2+ concentration from 1.0 to 0.05 mM did not modify SZ95 sebocyte numbers after 24 h compared with SZ95 sebocytes maintained at 1.4 mM ecCa2+ (Fig. 3).
A significant, ecCa2+ level-dependent decrease in neutral lipid content in SZ95 sebocytes was assessed with increasing ecCa2+ levels from 0.05 to 1.4 mM at 24 h (Fig. 4). On the other hand, a decrease in ecCa2+ levels from 1.4 to 0.05 mM was associated with an increase in neutral lipid content at ecCa2+ levels of 0.05 mM compared with that at ecCa2+ levels of 1.4 mM at 24 h (p < 0.05) (Fig. 4).
Fig. 4. Effect of ecCa2+ levels on SZ95 sebocyte neutral lipids at 24 h. The results presented are from 1 representative experiment out of 2, sixplicate ones performed and are shown as the correlation of neutral lipids/cell after measuring the absolute fluorescence units (AFU) of the correlating Nile red assay (NR) and fluorescein diacetate assay (FDA). *p < 0.05; **p < 0.01.
Caspase 3/7 activity was elevated at lower ecCa2+ levels in comparison with higher ones. The elevation was ecCa2+ level-dependent, being highest at 0.05 mM ecCa2+ (Fig. S1).
To detect the expression levels and activity of E-cadherin in SZ95 sebocytes under different ecCa2+ levels, the cells were cultured for 24 h under 0.05–1.4 mM ecCa2+ and 72 h under 0.05 and 1.4 mM ecCa2+. The icCa2+-dependent transmembrane glycoprotein E-cadherin was found to be ubiquitously expressed in SZ95 sebocytes independently of the ecCa2+ levels at 24 h, whereas the cells expressed minor increase in E-cadherin levels at 72 h under high ecCa2+ (Fig. 5a). However, its intercellular immunofluorescence signal was only present in subconfluent SZ95 sebocyte cultures under high ecCa2+ levels (Fig. 5b) and was almost lost in the few attached cells under low ecCa2+ levels (Fig. 5c).
Fig. 5. Effect of ecCa2+ levels on the expression and distribution of the icCa2+-dependent transmembrane glycoprotein E-cadherin in SZ95 sebocyte cultures. (a) Western blot analysis was used to monitor the expression levels of E-cadherin. Cell lysates were analysed after incubation with different concentrations of ecCa2+ for 2 different times. Thirty µg of protein each were separated by sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) (7%). β-actin was used to confirm similar protein loading across samples. (b, c) The distribution, and therefore activity of E-cadherin, in SZ95 sebocyte cultures maintained under high (1.4 mM; b) and low (0.05 mM; c) ecCa2+ levels was detected by immunofluorescence shown in green (blue represents nuclear counterstaining).
To study and reassure the effects of ultimately low ecCa2+ conditions on SZ95 sebocytes, cells were maintained at high ecCa2+ levels and treated for 24 h with the potent ecCa2+ chelating agent EGTA at concentrations of 0.05–0.4 mM. While cell numbers were not affected (except at EGTA 0.4 mM at which cells detached and floated in the medium), neutral, but not polar lipids significantly increased at EGTA concentrations ≥ 0.3 mM (p < 0.01) (Fig. 6).
Fig. 6. Effects of ecCa2+ chelation with EGTA (ethylene glycol-bis(β-aminoethyl ether)-N,N,N’,N’-tetraacetic acid) on SZ95 sebocyte numbers (dotted line) and lipogenesis (bars). The results presented are from 1 out of 3 representative experiments performed in sixplicate and are calculated as the absolute fluorescence units (AFU) of each experiment. *p < 0.05; **p < 0.01.
Moreover, SZ95 sebocytes showed a highly developed Golgi complex and increasing small lipid droplets consistent with an upregulation of lipid synthesis (Fig. 2e).
1,25(OH)2D3 did not affect the SZ95 sebocyte numbers until 72 h of treatment (Fig. S2). On the other hand, it decreased the amount of neutral lipids/cell at high pharmacological doses (0.1 µM), an effect which was independent of ecCa2+ levels (p < 0.05) (Fig. S3).
Moreover, 1,25(OH)2D3 induced autophagic vesicles in SZ95 sebocytes, which were at late differentiation stage, independently of the ecCa2+ levels, (Fig. 2 f, g). 1,25(OH)2D3 suppressed sebaceous lipogenesis, as seen by the reduction in the number of small intracellular lipid droplets.
The 104 patients with acne evaluated exhibited mild (n = 81), moderate (n = 22) and severe (n = 1) acne (Table SI). The total Ca2+ levels of all patients with acne and controls were within the normal limits (2.2–2.6 mM with 50% ionized Ca2+). Patients with acne presented a median ionized Ca2+ level of 1.21 mM (95% CI 1.19–1.22 mM), while the 112 control patients presented a median ionized Ca2+ level of 1.16 mM (95% CI 1.17–1.19 nM) (p < 0.0001). Patients with acne aged ≤ 24 years (n = 51) presented higher ionized Ca2+ levels (median 1.22 mM, 95% CI 1.22–1.24 mM) than both older patients with acne (n = 53; median 1.19 mM, 95% CI 1.18–1.20 mM) and controls (p = 0.001).
On the other hand, the patients with acne exhibited a median 1,25(OH)2D3 serum level of 0.31 nM (95% CI 0.26–0.36 nM). The 112 control patients presented a median 1,25(OH)2D3 serum level of 0.24 nM (95% CI 0.15–0.33 nM) (ns). Patients with acne aged ≤ 24 years presented lower 1,25(OH)2D3 serum levels (median 0.25 nM, 95% CI 0.21–0.29 nM) than older patients with acne (median 0.38 nM, 95% CI 0.30–0.46 nM) (p = 0.012).
The epidermis maintains a steep Ca2+ gradient, with the highest levels in the stratum granulosum and lowest levels in the stratum basale, in parallel with a gradient of Ca2+ receptor expression (30). This distribution of Ca2+ enables proliferation of keratinocytes in the basal epidermal layers, while promoting their differentiation in the upper ones. 1,25(OH)2D3 increases the icCa2+ concentration and increases the mRNA and protein levels of keratinocyte differentiation markers, in many cases in a synergistic manner with Ca2+ levels (31, 32).
Beyond keratinocytes, Ca2+ is being increasingly recognized as a central signal transmitter in other cells. Ca2+ is critical for cell function. The role of 1,25(OH)2D3 in controlling Ca2+ cell handling is a universal feature and explains the presence of VDR in practically all tissues and the fact that 1,25(OH)2D3 and its active metabolites regulate so many physiological functions not obviously involved in Ca2+ homeostasis (31, 33–35).
1,25(OH)2D3 has been shown to suppress lipogenesis in hamster sebaceous gland cells in vitro (36). To repeat the experiments on human material, the immortalized human sebaceous gland cell line SZ95, which was shown to maintain the major characteristics of normal sebocyte differentiation (27), has been used to gain first insight into the function of vitamin D endocrine system in human sebaceous glands. Incubation of SZ95 sebocytes with 1,25(OH)2D3 resulted in a dose-dependent suppression of lipogenesis (25, 37). Using real-time PCR, it was demonstrated that key components of the vitamin D system (VDR, vitamin D-25-hydroxylase (25OHase), 25-hydroxyvitamin D-1α-hydroxylase, and 1,25-dihydroxyvitamin D-24-hydroxylase) are strongly expressed in SZ95 sebocytes (25).
Despite the common embryologic origin of epidermal keratinocytes and sebocytes, we have previously reported a differential response of SZ95 sebocytes to ecCa2+ (15). In this study, 1,25(OH)2D3 exerted a differential effect on SZ95 sebocyte numbers depending on its concentration and on ecCa2+ levels. On the other hand, a reduction in lipid synthesis was found after the incubation of SZ95 sebocytes with 1,25(OH)2D3, which was independent of ecCa2+ concentrations. 1,25(OH)2D3 may inhibit sebaceous lipogenesis by binding VDR/RXR dimers, a well-studied pathway also active in retinoid biology, where RAR/RXR dimers are regulated (25). On the other hand, high ecCa2+ upregulates terminal keratinocyte differentiation of epithelial cells, a pathway that has been shown to inhibit sebaceous differentiation and thus lipogenesis (23).
In human keratinocytes, icCa2+ has been recognized as an important player in the modulation of Ca2+ signalling and a major regulator of keratinocyte biology (1, 38–41). In contrast, the downstream signalling pathway by which ecCa2+/icCa2+ and 1,25(OH)2D3 exert their function on SZ95 sebocytes had not been elucidated until now. In sebocytes, α-melanocyte-stimulating hormone was shown to control Ca2+ response by increasing icCa2+ sequestering (20). In agreement with that, α-melanocyte-stimulating hormone increased lipid synthesis in SZ95 sebocytes (19). SZ95 sebocytes express transient receptor potential vanilloid-1, a non-selective, Ca2+-permeable channel, that, when stimulated, selectively modulates lipogenesis (20, 37). These finding indicate that there may be a correlation and active communication regarding the Ca2+ levels between the 2 compartments. Interestingly, the expression of the transmembrane glycoprotein E-cadherin is icCa2+-dependent (42, 43). Our findings indicate that reduction in ecCa2+ levels induces differentiation, lipogenesis and apoptosis of human sebocytes (16). On the other hand, SZ95 sebocyte icCa2+ levels seem to be strongly influenced by ecCa2+ levels up to 6 days in culture: although E-cadherin expression levels did not markedly change, its activity was strongly reduced, since its transmembrane distribution almost vanished and cells became dissociated. The transmembrane distribution is a major indicator of active E-cadherin and the reduction in this distribution and the subsequent cell dissociation is a marker of E-cadherin inactivation (44). Moreover, 1,25(OH)2D3 partially inhibited sebaceous lipogenesis, probably by regulating the icCa2+ gradient inducing autophagic vesicles in the cells. These two events indicate that 1,25(OH)2D3 can induce lipophagy in human sebocytes, which is a cell mechanism for intracellular generation of energetically efficient essential components, such as free fatty acids and sugars (45).
These interesting biological effects may indicate that ionized Ca2+ and 1,25(OH)2D3 are likely to affect human sebocytes in vivo. In vivo effects on sebocyte proliferation and lipogenesis, like the ones detected in the present work, may occur by ionized Ca2+ serum levels and/or 1,25(OH)2D3 insufficiency or after systemic supplementation (46). The increased ionized Ca2+ levels detected in the young patients with acne in our clinical study are compatible with the increased sebocyte numbers and reduced apoptosis, which were detected with higher ecCa2+ concentrations in our in vitro study and the enhanced sebocyte proliferation observed in acne-involved glands (47). Interestingly, sebocyte hyperactivity is induced by androgens (48) and reduced apoptosis is detected under peroxisome proliferator-activated receptor activation (49), both being major inducing factors of acne pathogenesis (50). However, the very narrow concentration levels in vivo may not allow the complete display of Ca2+ effects on the sebaceous gland in vivo. On the other hand and although 1,25(OH)2D3 serum levels did not differ among patients and controls, young patients with acne exhibited lower 1,25(OH)2D3 serum levels than older ones, which may partially explain the hyperseborrheic status of young patients with acne.
In conclusion, ecCa2+ levels and 1,25(OH)2D3 were shown to exert a regulatory function on human sebocytes in vitro, including an autophagy/lipophagy effect in SZ95 sebocytes induced by 1,25(OH)2D3 and an ecCa2+-controlled, previously not reported, endogenously active channel mechanism (51). Both agents, but mostly 1,25(OH)2D3 levels, may affect sebaceous lipogenesis in vivo. The increased ionized calcium as well as the reduced 1,25(OH)2D3 levels detected in the serum of younger patients with acne may contribute to the increased sebaceous gland volume and the seborrhea, which are characteristic acne signs.
G.K. Menon was partially supported by Avon Products, Inc. in the initial steps of this study. H. Seltmann has been supported by grants of the Deutsche Forschungsgemeinschaft (DFG), the Bundesinstitut für gesundheitlichen Verbraucherschutz und Veterinärmedizin (BgVV-ZEBET) and the Bundesministerium für Bildung und Forschung (BMBF) to C. C. Zouboulis.
The authors declare no conflicts of interest.


 Click to show fullsize
Click to show fullsize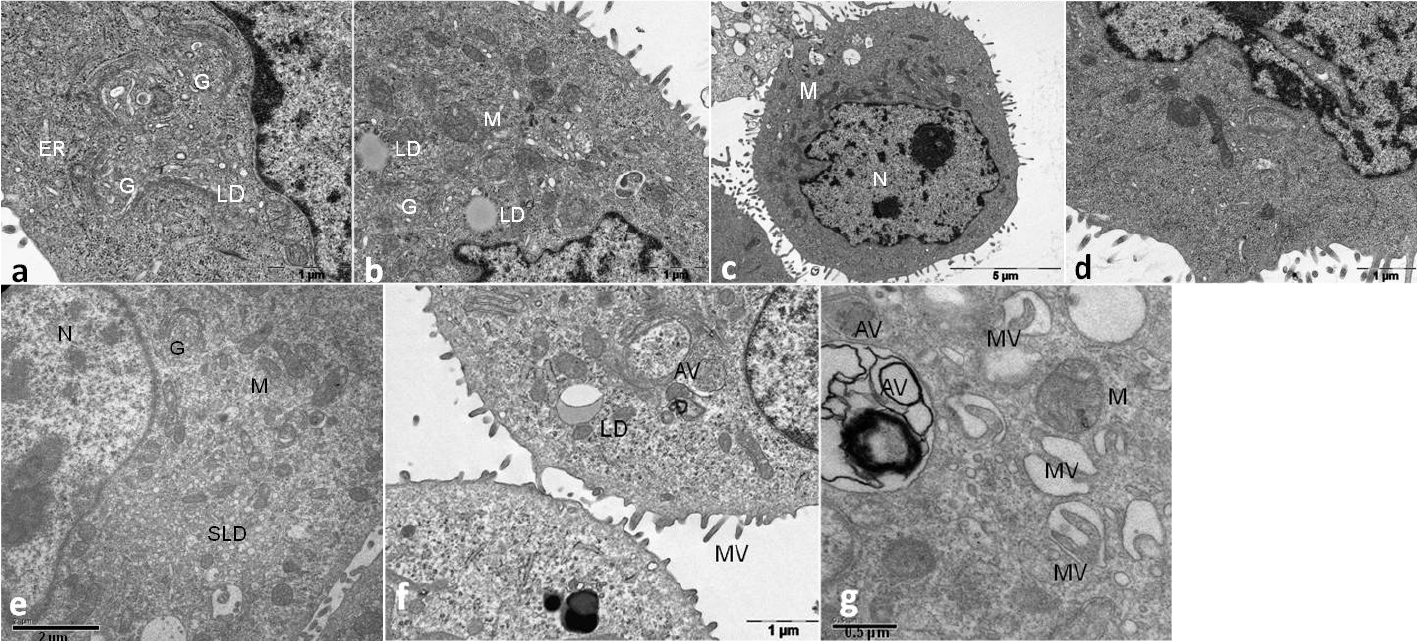 Click to show fullsize
Click to show fullsize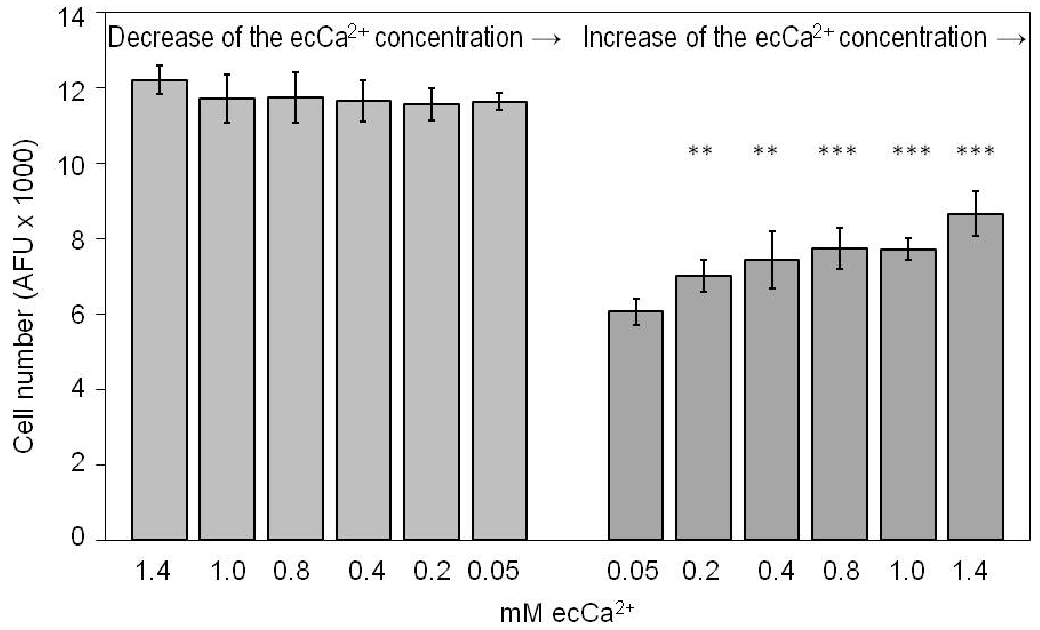 Click to show fullsize
Click to show fullsize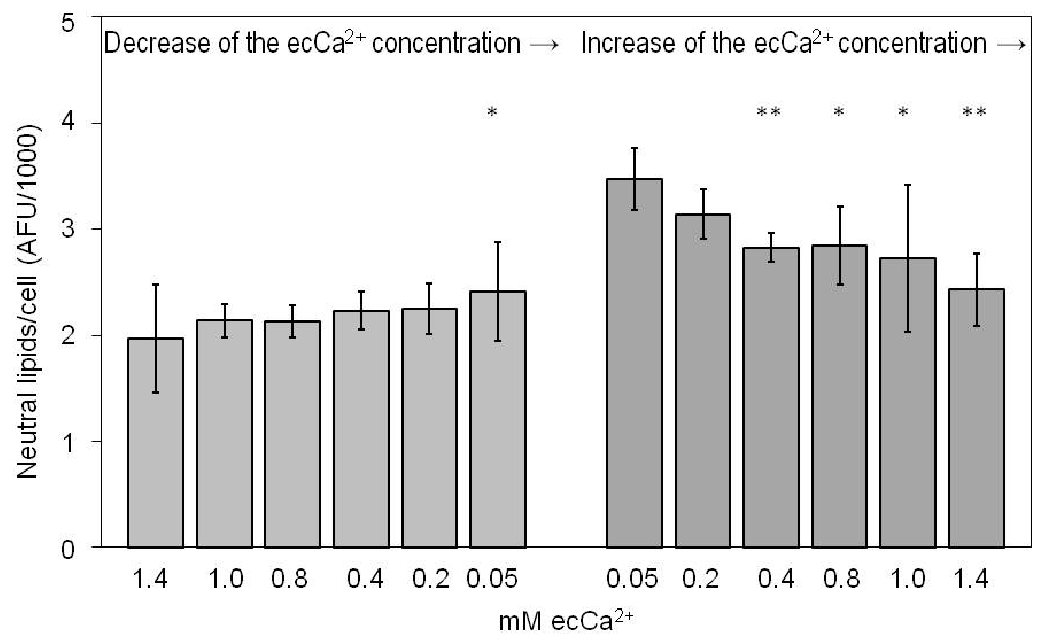 Click to show fullsize
Click to show fullsize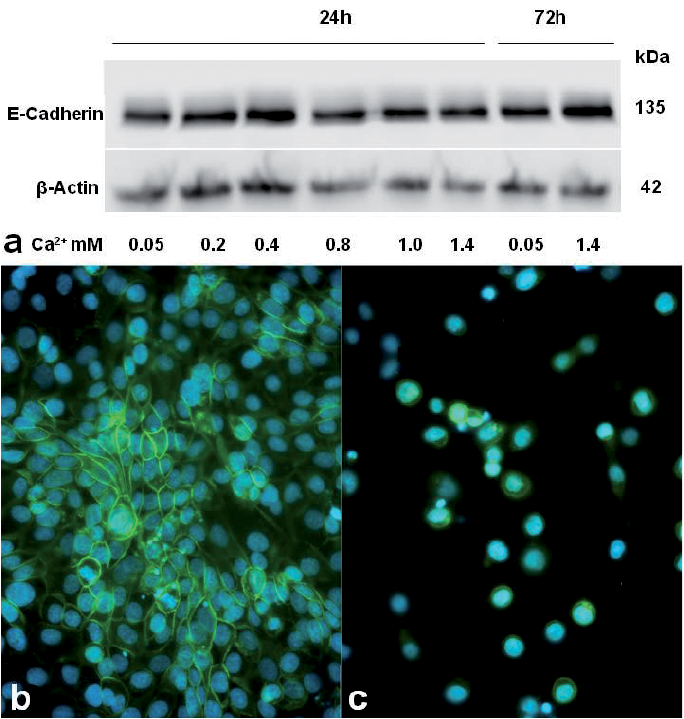 Click to show fullsize
Click to show fullsize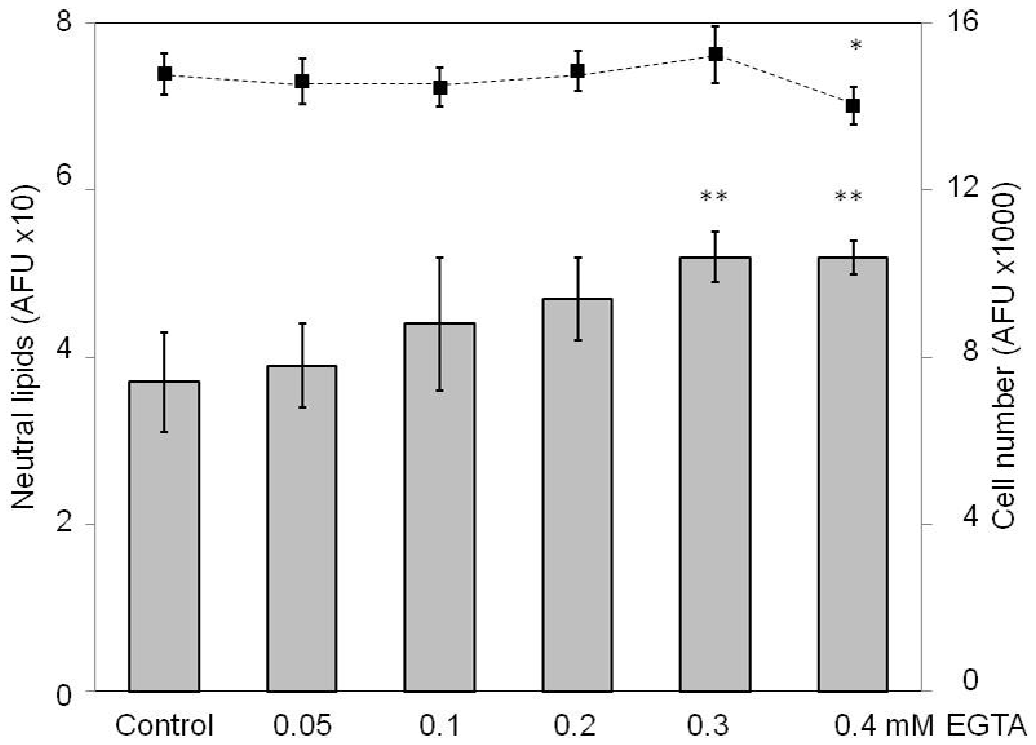 Click to show fullsize
Click to show fullsize