


1Department of Odontology, 2Department of Clinical Medicine and Public Health, Dermatology and Venerology, and 3Department of Medical Biosciences, Umeå University, Umeå, Sweden
Lichen planus is a chronic recurrent inflammatory disease affecting both skin and mucosa, mainly in oral and/or genital regions. Keratinocytes go through a well-regulated process of proliferation and differentiation, alterations in which may result in defects in the protective epithelial barrier. Long-term barrier impairment might lead to chronic inflammation. In order to broaden our understanding of the differentiation process in mucosal lichen planus, we mapped the expression of 4 factors known to be involved in differentiation. Biopsies were collected from oral and genital lichen planus lesions and normal controls. Altered expression of all 4 factors in epithelium from lichen planus lesions was found, clearly indicating disturbed epithelial differentiation in lichen planus lesions.
Key words: lichen planus; differentiation; epithelial barrier.
Accepted Sep 7, 2016; Epub ahead of print Sep 7, 2016
Acta Derm Venereol 2017; 97: XX–XX.
Karin Danielsson, Department of Odontology, Umeå University, SE-901 85 Umeå, Sweden. E-mail: karin.p.danielsson@umu.se
Lichen planus (LP) is a chronic recurrent inflammatory disease affecting both skin and mucosa, mainly in oral and/or genital areas. The subgroup oral lichen planus (OLP) affects approximately 2% of the population in Sweden (1). Many patients with OLP also have skin and/or genital lesions. A recent study showed 45% of men and 53% of women to have oral as well as genital lesions (2). The aetiology of LP is unknown, but autoimmunity has been suggested to play a part in its pathobiology, and autoantibodies against factors involved in epithelial homeostasis, such as members of the p53 (3) family and ELF-3 (4) have been found in sera from patients with OLP. Current treatment is unsatisfactory and many patients respond poorly or not at all to the first choice of treatment, topical corticosteroids.
Skin and mucosa exert a barrier and a first line of defence, protecting against invasion of microorganisms and allergens, and the epithelium is also an essential part of the immune system. Keratinocytes go through a well-regulated process of proliferation and differentiation, and alterations in this process may result in defects in the protective epithelial barrier. It has been suggested that long-term barrier impairment can lead to chronic inflammation (5). Previously we have shown altered expression of p63, which is important in maintaining epithelial homeostasis, and ELF-3, which is involved in differentiation, in OLP (3, 4). A whole-genome expression study performed on micro-dissected epi-thelium from LP, oral and genital, and corresponding healthy controls further showed that the biological processes most affected in LP were those involved in differentiation, keratinization and development of the epithelium (6).
In order to broaden our understanding of the differentiation process in LP, we mapped the expression of 4 factors known to be involved in differentiation and to be altered in different types of cancers and inflammatory disorders, but not previously associated with LP lesions. These factors were chosen from the gene list generated by whole-genome array analysis (6), and analysed both at RNA and protein level.
First we evaluated the expression of Stratifin (SFN) or 14-3-3σ, which is specifically expressed in stratified squamous cells. SFN has many roles and can act as a tumour suppressor, a cell cycle checkpoint regulator and a regulator of epithelial homeostasis. SFN is involved in epithelial differentiation in skin (7) and increased levels are seen in differentiating keratinocytes after DNA damage stress (8). SFN is also important for the formation of desmosomes in squamous epithelial cells (9). In hyperproliferative disorders, such as actinic keratosis, psoriasis and squamous cell carcinoma, higher expression of SFN is seen (10), in contrast to the lower expression seen in oesophageal cancer (11).
Epithelial membrane protein 1 (EMP1) is a member of the PMP22 family that was first described in 1995. It can be detected in epithelial tissue from the gastrointestinal tract, skin, lung and brain, and is involved in regulating proliferation and differentiation of epithelial cells (12). Recently EMP1 was reported as an important player in the formation of tight junctions (13). Alteration of EMP1 expression has been implicated in many types of cancer, and in head and neck squamous cell carcinoma and oesophageal cancer down-regulation is seen. It has also been suggested that EMP1 is associated with lymph node metastasis in oral squamous cell carcinoma (14). In nasal polyps with chronic inflammation, hyperplasia and metaplasia, down-regulation of EMP1 has been noted (15).
Inhibitor of DNA-binding 3 (ID3) belongs to a large family of helix loop helix (HLH) transcription factors and is 1 of the 4 members of the HLH class V subfamily. The ID proteins act as negative regulators of transcription. In squamous epithelia, expression of ID3 is found in the proliferating basal layers and levels decrease with increased differentiation. High expression of ID3 is related to an undifferentiated, proliferative keratinocyte phenotype and high levels of ID3 have been found in poorly differentiated SCC (16). ID3 also has the ability to regulate differentiation of CD8+ T cells (17).
The 4th factor, Kazrin (KAZN), has the ability to regulate adhesion and differentiation of keratinocytes and can bind to periplakin, a protein found in the epidermal cornified envelope, localized to desmosomes (18). KAZN is up-regulated during keratinocyte differentiation, and overexpression in keratinocytes leads to changes in morphology and impaired assembly of intercellular junctions. KAZN is expressed in all layers of stratified squamous epithelia (19).
Altered expression of all 4 factors was found in the epithelium from LP lesions, clearly showing a disturbance in factors involved in epithelial differentiation in LP lesions.
From 16 patients clinically and histologically diagnosed with LP in the oral and/or genital mucosa, 21 punch biopsies (12 from oral lesions and 9 from genital lesions) were collected. From 5 patients biopsies were collected from both OLP and genital LP (GLP). Four patients were men (mean age 58 years, range 45–67 years) and 12 women (mean age 66 years, range 45–77 years). OLP was diagnosed according to the modified WHO diagnostic criteria (20). Control biopsies were collected from 8 persons with healthy oral (n = 8) and genital (n = 5) mucosa. Five of these control biopsies were taken from oral and genital mucosa in the same individual. Two of the oral control persons were men aged 49 and 69 years, and 6 were women with a mean age of 55 years, range 45–69 years. The genital control biopsies were all from women (n = 5) with a mean age of 55 years, range 45–67 years (Table SI). Biopsies were embedded in Tissue TEK, snap-frozen in liquid nitrogen and stored at –80ºC until use.
For the immunohistochemical (IHC) analysis formalin-fixed and paraffin-embedded (FFPE) samples, 22 OLP, 19 GLP (Table SI) and 15 healthy oral and 7 healthy genital controls were retrieved from the archives in the Department of Clinical Pathology, University Hospital, Umeå, Sweden. Of the 19 OLP patients 17 were women (mean age 60 years, range 42–85 years) and 5 men (mean age 43 years, range 34–70 years). In the GLP group 13 were women (mean age 61 years, range 40–82 years) and 6 men (mean age 45 years, range 33–70 years).
The study was approved by the local ethics review board at Umeå University (Dnr 09-083M).
Ten µm cryosections were placed on membrane-coated glass slides (Carl Zeiss Micro Imaging GmbH, Bernried, Germany) and stained with HistoGene staining solution (Arcturus Bioscience, Mountain View, CA, USA). Laser micro-dissection was performed using PALM® micro laser system (PALM GmbH, Germany), and dissected epithelium collected and placed in tubes with 850 µl TRIzol Reagent (Invitrogen). Tubes were incubated at room temperature for 30 min and then mixed for 5 min before centrifugation. Tubes were stored at – 80°C until RNA extraction.
Samples were thawed on ice and mixed for 1 min, followed by spinning at 12,000 g for 10 min at +4°C. The supernatant was transferred to a new tube and 160 µl chloroform added. Samples were mixed for 15 s then incubated at room temperature for 3 min followed by centrifugation at 12,000 g for 15 min at +4°C. The aqueous phase was collected and transferred to a new tube. RNeasy Plus Microkit, Qiagen (Hilden, Germany) was then used for purification of total RNA.
For cDNA synthesis 200 ng of RNA and the RevertAid H Minus First Strand cDNA Synthesis Kit (Fermentas, St Leon-Rot, Germany) was used. For detection of mRNA levels of SFN, ID3, EMP1 and KAZ qRT-PCR was used. iQ™ SYBR® Green Supermix (Bio-Rad Laboratories Inc., Hercules, CA, USA) and QuantiTect Primer Assay® (Qiagen) was used for PCR amplification. All samples were run in duplicate and normalized against GAPDH (Qiagen). Statistical analysis was performed using IBM SPSS Statistics 20 and the non-parametric method Mann-Whitney. The significance level was set at p < 0.05.
Five µm sections were cut from formalin-fixed paraffin-embedded samples from 19 OLP and 19 GLP and 15 oral and 7 genital controls. The antibodies used were for SFN, anti 14-3-3 sigma antibody ab 14123 (Abcam, Cambridge, UK) diluted 1:100, for ID3, anti ID3 antibody ab 118032 (Abcam) diluted 1:50, for EMP1, anti EMP1 HPA056250 (Sigma Aldrich, Sweden AB, Stockholm, Sweden) diluted 1:50 and for KAZN, anti KAZN HPA032096 (Sigma Aldrich) diluted 1:25. Staining was performed in a Ventana Bench Mark Ultra (Ventana Medical Systems, Inc., Tucson, AZ, USA) staining machine according to the manufacturer’s recommendations.
Staining was evaluated independently by 2 of the authors (KD, KN) using the so-called Quick score (QS) method (21). The proportion of epithelial cells stained and the intensity of staining was evaluated. The proportion of cells stained was classified from 1 to 6 (1: 0–4%, 2: 5–19%, 3: 20–39%, 4: 40–59%, 5: 60–79% and 6: 80–100%). For intensity, 4 levels were used (0: negative, 1: weak, 2: intermediate, 3: strong). Scores from staining and intensity for each slide were multiplied, giving a QS. Results from the scoring were compared and cases of disagreement discussed until consensus was reached.
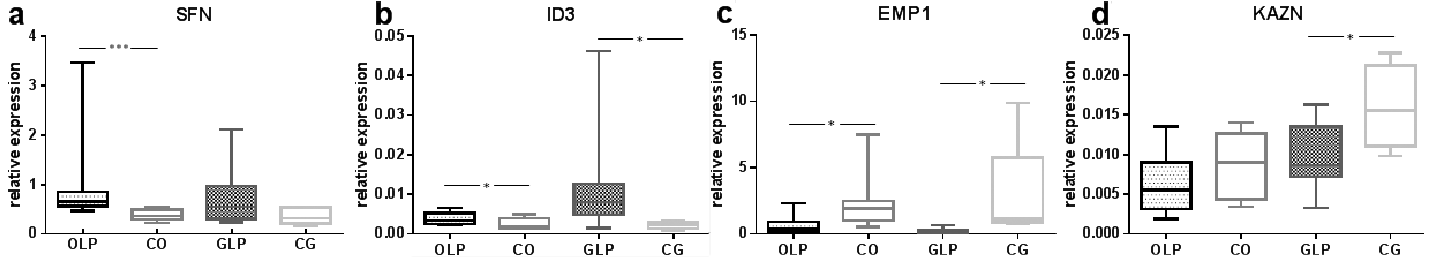
Expression of SFN, EMP1, ID3 and KAZN was analysed using quantitative RT-PCR mRNA. Higher expression of SFN (Fig. 1a) and ID3 (Fig. 1b) was found in the epithelium of both OLP and GLP lesions compared with controls. For SFN there was a significant difference (p ≤ 0.001) between OLP and controls, while the difference between GLP and controls was not significant (p ≤ 0.178) (Fig. 1a). Expression of ID3 was significantly higher in both OLP (p ≤ 0.045) and GLP (p ≤ 0.014) compared with controls (Fig. 1b).
Fig. 1. PCR showing significantly higher expression of: (a) Stratifin (SFN) and (b) Inhibitor of DNA-binding 3 (ID3) mRNA in both oral lichen planus (OLP) and genital lichen planus (GLP) compared with normal. Expression of (c) Epithelial membrane protein 1 (EMP1) and (d) Kazrin (KAZN) were both decreased in OLP and GLP compared with normal. *p < 0.05, ***p < 0.005. CO: control oral; CG: control genital.
For EMP1 and KAZN expression was lower in both OLP and GLP compared with controls. EMP1 expression was significantly lower in both OLP (p ≤ 0.045) and GLP (p ≤ 0.014) (Fig. 1c), while expression of KAZN was significantly lower only in GLP (p ≤ 0.042) (Fig. 1d).
Examples of stainings are shown in Fig. 2. Quick scores (QS) ranged from 0 to 18 for SFN, between 3 and 18 for ID3 and between 1 and 12 for KAZN, whereas for EMP1 QS did not exceed 6 (Table SII). Due to insufficient amount of material and artefacts in staining, it was not possible to evaluate all slides for all 4 factors.
Fig. 2. Immunohistochemistry (IHC) staining (× 20) showing examples of expression of Epithelial membrane protein 1 (EMP1), Inhibitor of DNA-binding 3 (ID3), Kazrin (KAZN) and Stratifin (SFN) in control oral (CO), oral lichen planus (OLP), control genital (CG) and genital lichen planus (GLP).
For SFN a significant difference was seen between OLP and normal oral mucosa, with higher expression in OLP (p ≤ 0.001). In OLP 83% had a QS ≥ 10, whereas in normal oral mucosa only 7% had a QS of 10 or more. For GLP no significant difference was seen compared with its normal counterpart, even if the number of high expressing samples with a QS ≥ 10 was higher in GLP; 74% compared with 67% in controls (Table SII).
For ID3 when looking at protein levels all genital control samples had a QS of 10 or above, compared with 58% of the GLP cases. For OLP the majority of samples (68%) had high QS compared with only 40% of the oral controls (Table SII).
For KAZN, normal genital mucosa had higher protein levels compared with GLP. None of the genital control samples had a QS below 10. OLP samples showed more high expressing samples (71%) compared with controls (53%) (Table SII).
EMP1 expression did not exceed a QS of 6 for normal or for LP mucosa. Seventy-four percent of OLP samples and 53% of the normal oral samples were negative, showing a QS of 0. In GLP 37% were negative, whereas none of the normal genital controls were negative (Table SII).
Differentiation is a process that has previously been shown to be dysregulated in LP lesions (6). The present study examined whether factors with a known role in this process in general, but not previously connected to LP lesions in particular, also are dysregulated in this disease. Four genes were chosen from the gene list generated by whole-genome array analysis, 2 up-regulated and 2 down-regulated (6), and analysed their expression at both mRNA and protein levels in OLP and GLP lesions compared with their respective normal control tissues.
Even if the same samples were not used in the mRNA and protein analyses there was, in general, good concordance between levels of mRNA and protein. There were 2 exceptions to this concordance, where the significant up-regulation of mRNA levels of ID3 in GLP samples compared with controls not could be seen at protein levels, and the down-regulation, though not significant, in mRNA levels of KAZN in OLP samples compared with controls not was seen at protein levels. A possible explanation is that expression levels of mRNA for both ID3 and KAZN were very low; up to 1,000 times lower than for SFN and EMP1, which both showed good concordance between mRNA and protein levels.
For SFN and EMP1 OLP lesions showed significantly higher levels of SFN and significantly lower levels of EMP1 compared with controls. GLP showed the same pattern even if levels of SFN not were significantly higher compared with genital controls. For protein levels results were comparable, as both OLP and GLP showed higher expressing samples compared with controls. Increased expression of SFN has previously been shown in differentiating keratinocytes (8). A side-effect of SFN overexpression was seen in mice as a decrease in epithelial thickness, most likely due to impaired proliferation of keratinocytes (22). The increase in SFN seen in OLP and GLP could be a way of stimulating differentiation in lesional keratinocytes, but as a side-effect these increased levels give rise to the commonly seen atrophic epithelium. SFN affects collagen and hyaluronan metabolism as well as the barrier function of skin and mucosa, and increased levels affect water-holding capacity and barrier function (23).
The lower expression of EMP1 seen in both OLP and GLP also affects differentiation, rendering less differentiated keratinocytes, a disturbed function of adherence and tight junctions and impaired epithelial barrier, as has been seen in respiratory epithelium (24). Lower expression of EMP1 could thus lead to impaired barrier function and influence both adherence and tight junction formation in LP epithelium.
Expression of ID3, on the other hand, is generally high in proliferating, undifferentiated cells and down-regulated during differentiation (25). The higher expression of ID3 seen in both OLP and GLP epithelium could thus indicate higher proliferation. The increased expression of ID3 is in accordance with other studies showing increased expression of ID3 in inflammatory conditions (24). It is also known that ID proteins have many functions in lymphocyte development and are also predominantly expressed in lymphocytes (17). The higher expression of ID3 noted in both OLP and GLP epithelium could be caused by the high amount of lymphocytes present in the underlying connective tissue and also found within the epithelium. Decreased expression of KAZN mRNA was seen in both types of LP, but significantly changed only in GLP (p ≤ 0.042). When it comes to protein expression an increase was seen in OLP, while a slight decrease was seen in GLP. KAZN can induce terminal differentiation of keratinocytes and decreased expression leads to increased proliferation (19). Sevilla et al. found KAZN to be a regulator of intercellular adhesion, where high levels disrupt cell-cell adhesion, thus facilitating cells moving from basal to suprabasal layers (19). The increased expression of KAZN points towards an epithelium more terminally differentiated, at least in OLP. Even if results are not concordant they show deregulated expression of KAZN in both OLP and GLP compared with controls.
Taken together, the present analysis of GLP and OLP lesions show altered expression of factors involved in differentiation, but previously not connected with LP. Importantly, OLP and GLP show the same changes, supporting the classification of LP as a generalized disease with different sublocations. Exactly how each individual factor affects keratinocyte differentiation in LP lesions is not clear, but all 4 factors affect the process in some way. The altered levels of SFN, EMP1 and KAZN furthermore indicate impaired barrier function with disturbances in desmosomes, adherens and tight junctions. With regard to therapeutic possibilities, these novel findings pinpoint factors causing deregulated differentiation as possible novel targets.
This study was supported by grants from the Medical Faculty, Umeå University, Västerbotten County Council and Lion’s Cancer Research Foundation.
The authors declare no conflicts of interest.


 Click to show fullsize
Click to show fullsize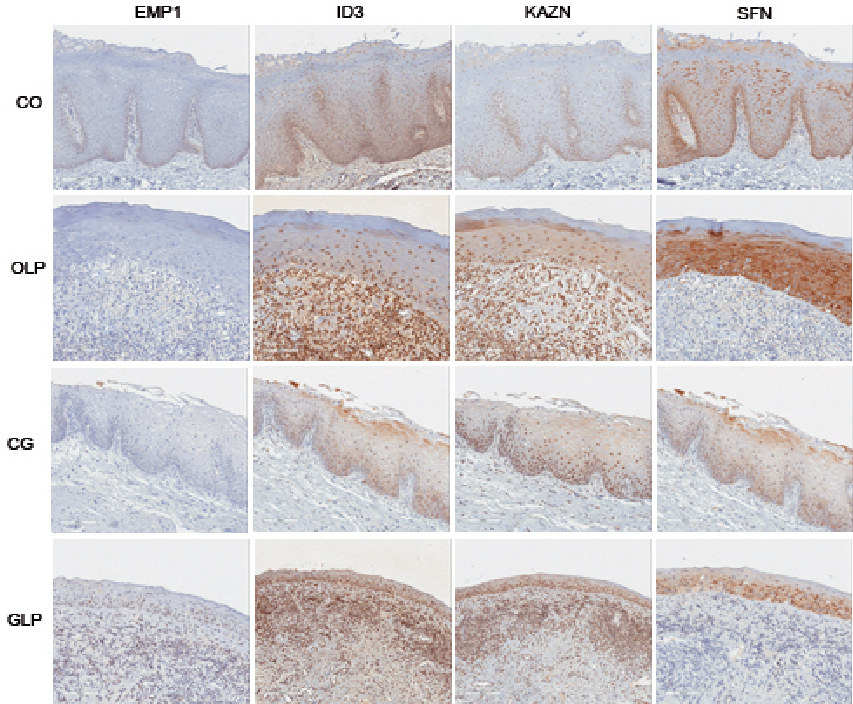 Click to show fullsize
Click to show fullsize