


1PEDEGO Research Unit, Oulu Center for Cell-Matrix Research, Department of Dermatology, Medical Research Center Oulu, Oulu University Hospital and University of Oulu, 2Cancer and Translational Medicine Research Unit, Department of Pathology, Medical Research Center Oulu, Oulu University Hospital and University of Oulu, and 3Biomedicine, Medical Microbiology and Immunology Research Unit, University of Oulu, Oulu, Finland
The effects of topical calcipotriol/betamethasone combination therapy and betamethasone monotherapy on inflammatory T-cell numbers and molecular markers were compared in patients with psoriasis. Combination therapy down-regulated the expression of tumour necrosis factor (TNF)-α, interleukin (IL)-23A, IL-17A, S100A7, CCL-20 and interferon (IFN)-γ in skin and TNF-α, IL-6, IL-23A, T-bet and IFN-γ in peripheral blood mononuclear cells (PBMCs). Betamethasone monotherapy had less effect. Expression of FoxP3 in both skin and PBMCs was down-regulated by calcipotriol/betamethasone, but not by betamethasone. Immunohistochemical analysis revealed that calcipotriol/betamethasone reduced the numbers of CD4+ and CD8+ T cells and Tregs in psoriatic lesions more than betamethasone. Flow cytometric analyses demonstrated that calcipotriol/betamethasone decreased the numbers of circulating CD8+ T cells, Tregs, skin-homing Th17 memory cells and Th22 memory cells, while betamethasone had little or no effect. Glucocorticoid receptors GRα and GRß were expressed in psoriatic skin. In conclusion, calcipotriol increases the immunosuppressive power of betamethasone by suppressing the inflammatory TNF-α – IL-23 – IL-17 axis.
Key words: glucocorticoid receptor; Treg; antimicrobial peptide; CCL-20; skin-homing receptor.
Accepted Nov 17, 2016; Epub ahead of print Nov 21, 2016
Acta Derm Venereol 2017; 97: XX–XX.
Corr: Minna Kubin, Department of Dermatology, Oulu University Hospital, Kajaanintie 50, FIN-90029 Oulu, Finland. E-mail: minna.kubin@ppshp.fi
A growing body of evidence indicates that different components of the innate and adaptive immune system are involved in initiating and maintaining the inflammatory response in psoriasis (1–4). Human genome-wide association studies and clinical studies have demonstrated up-regulation of the interleukin-17 (IL-17) pathway in psoriasis (2, 5, 6). In addition, dysfunction of regulatory T cells (Tregs) has been described (7), yet their role in psoriasis has been little-studied (8).
Topical therapies based on glucocorticoids (GC) and calcipotriol are usually sufficient to manage mild and moderate psoriasis (9). GCs produce their well-known anti-inflammatory effects through GC receptors (GR) and by controlling specific target genes and inhibiting several cytokines (10). Calcipotriol mediates its effects via binding to the nuclear vitamin D3 receptor (11). The anti-inflammatory effects of calcipotriol are inferior compared with those of GCs, but an incremental effect is seen when the 2 are combined (12). A common treatment for psoriasis is an ointment containing calcipotriol and betamethasone dipropionate (3, 6, 9). Several studies have demonstrated its effectiveness, superior tolerability and more rapid onset of action compared with those of its individual components (13–15). The in vivo effects of topical calcipotriol and betamethasone on Th17 cytokine expression in psoriasis have only been studied by Hino et al. (16): they observed a decrease in the frequency of circulating Th17 cells during 4 weeks therapy with calcipotriol and betamethasone. Calcipotriol monotherapy has been studied more widely (17, 18).
The aims of the current study were to investigate the effects of topically administered calcipotriol/betamethasone treatment on the expression of several cytokines and chemokines, and on the numbers of selected T-cell subsets in vivo. Furthermore, these effects were compared with betamethasone monotherapy. The effects of topical betamethasone on the expression of GR isoforms α (GRα) and β (GRβ) were also investigated. A further aim was to investigate whether there is a similar link between Th17 cytokines and up-regulation of GRβ expression in psoriasis that has been previously found in asthmatic patients (19).
The study was approved by the ethics committee of the Northern Ostrobothnia Hospital District of Finland, and performed according to the principles of the Declaration of Helsinki. Written consent was received from all participants.
Ten patients who had been referred to the Oulu University Hospital for mild to moderate plaque psoriasis were recruited to the study. The control group comprised 5 patients who had been referred for benign skin tumours.
Ten psoriatic patients were randomly assigned in a 1:1 ratio to receive topical treatment for one week with either a combination of 50 µg/g calcipotriol and 0.5 mg/g betamethasone dipropionate (Daivobet®; Leo Pharma, Vantaa, Finland) or betamethasone valerate ointment monotherapy (1 mg/g; Bemetson®; Orion Pharma, Espoo, Finland). Betamethasone valerate monotherapy was chosen over betamethasone dipropionate, since the latter is sold only in gel form in Finland. The patients applied the treatment once a day and the tubes were weighed. Controls received no treatment. Two 6-mm punch biopsies were taken from the patients before entering the study; one from untreated lesional skin and one from healthy skin. Another lesional 6-mm punch biopsy was taken after the treatment period from a symmetric contralateral plaque. Blood samples were taken before and after the treatment period. Peripheral blood mononuclear cells (PBMC) were isolated from heparinized blood using the Ficoll-Paque Plus density-gradient method (GE Healthcare Biosciences, Uppsala, Sweden). The cells were deep-frozen (–70°C) until further use. Psoriasis Area and Severity Index (PASI) (20) was measured by a sole, non-blinded observer (M.K.) at baseline and after the treatment period. A 6-mm skin punch biopsy was obtained from an area of healthy truncal skin in each of the controls. One half of each biopsy was fixed in formalin and the other was snap-frozen in liquid nitrogen for RNA extraction. Body mass index (BMI) was measured at baseline.
Total RNA was isolated from PBMCs with the QIAamp RNA Blood Mini Kit (Qiagen, Crawley, UK) and from skin biopsies with the RNeasy Plus Universal Mini kit (Qiagen). Reverse transcription was done using RevertAid reverse transcriptase (Thermo Fisher Scientific, Waltham, MA, USA). Quantitative real-time PCR (RT-qPCR) was performed as described previously (21) to quantify IL-6, IL-17A, IL-23A, interferon (IFN)-γ, tumour necrosis factor (TNF)-α, T-box expressed in T cells (T-bet, TBX21), forkhead/winged helix transcription factor 3 (FoxP3), S100A7, chemokine ligand (CCL)-20 and glucocorticoid receptor GRα and GRβ transcripts. Human glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and human β-actin served as controls. Normalized expression levels were compared with expression levels in normal skin/PBMCs. The primers are listed in Table SI.
Immunohistochemical staining of skin biopsies and digital image analysis with CD3, CD4, CD8, FoxP3, IL-17A positive cells and GR isoforms was performed as earlier described (21, 22). The antibodies are listed in Table SII. A negative control was operated similarly, but without the primary antibody. The images were captured from the area of each sample that represented the sample’s mean immune cell infiltration. Image analysis was performed using Java-based open-source image-processing software ImageJ version 1.45r (NIH, Bethesda, MA, USA) and a cell counting method (23) to record the mean number of positively stained cells per 20× field image (0.14 mm2). Data are presented as interquartile ranges (median positive cell count).
Frozen PBMCs were used for T-cell immunophenotyping. Memory CD4+ Th17/Th1 cells were analysed using a 6-colour flow cytometry (FCM) panel with BioLegend (San Diego, CA, USA) monoclonal antibodies (mAbs) against the surface antigens CD4-PerCP (344624), CD45RA-FITC (304106), CXCR3-APC (353708), CCR6-BV421 (353408), CCR10-PE (341504) and CLA-PECy7 (321316) following the protocol described by Ma et al. (24). For Treg analysis, anti-CD4-PerCP (345770), anti-CD25-APC (555434), and anti-CD127-PE (557938) mAbs from Becton Dickinson (Mountain View, CA, USA) were used for surface staining and anti-FoxP3-AF488 mAb (BioLegend; 320112) for intracellular staining according to the FoxP3/Transcription Factor Staining Buffer protocol (eBioscience, San Diego, CA, USA). CD4+ and CD8+ cells were verified from the lymphocyte population by staining with anti-CD4 -PerCP (344624; BioLegend) and anti-CD8-FITC(345772; Becton Dickinson). Samples were analysed with LSRFortessa FCM (BD Biosciences, Mountain View, CA, USA) and FlowJo software (TreeStar Inc, Ashland, OR, USA) (25).
Statistical analyses were conducted using IBM SPSS Statistics 22.0 (IBM, Chicago, IL, USA). Wilcoxon signed-rank and Mann–Whitney tests were used to analyse the results of immunohistochemical stainings. The paired sample t-test was used to compare RT-qPCR and FCM results before and after the treatment period. Spearman’s rho was used to analyse correlations between variables.
After one week’s treatment, PASI scores decreased in all 10 patients. The change in score was more marked (p = 0.346) in patients treated with combination therapy. Two of the patients treated with monotherapy showed only a minor effect: their PASI scores did not change markedly. The median BMI at baseline was 24, 26.5 and 26.7 in the combination therapy, monotherapy and control groups, respectively. The median amount of ointment used did not differ markedly between the therapies (Table SIII).
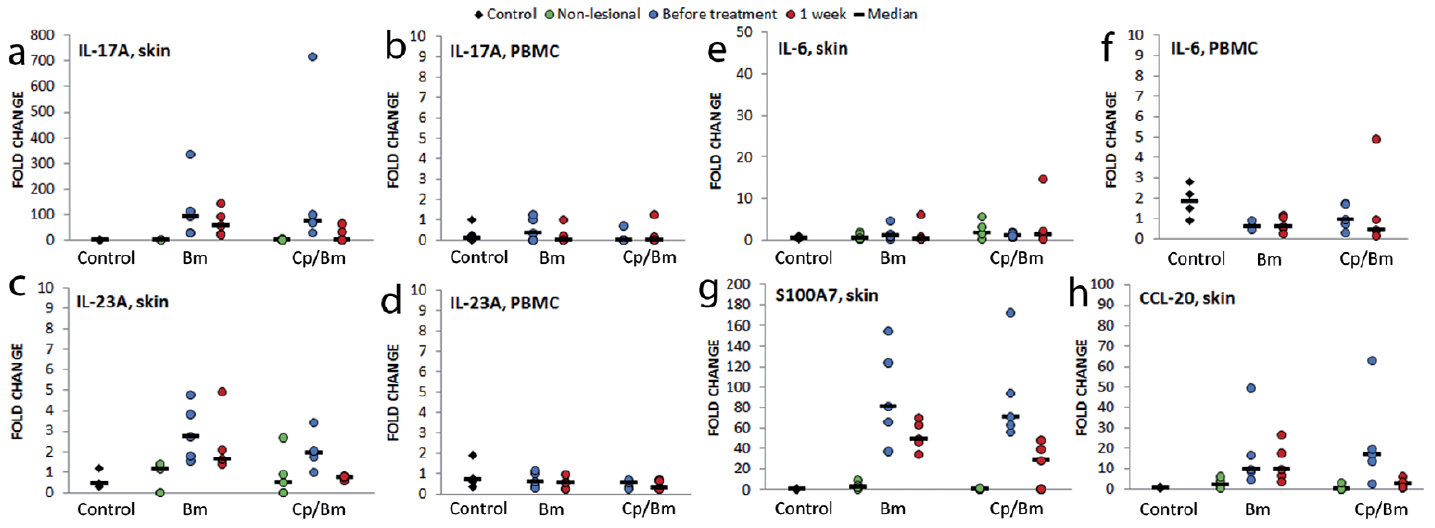
In each skin sample taken from both psoriatic patients and controls, only a few cells stained positively for anti-IL-17A in the epidermis (Table SIV, Fig. S1a). There were significantly more IL-17A-positive dermal cells in psoriatic lesional than in non-lesional (p = 0.004) or control (p = 0.036) skin samples (Table SIV, Fig. S1a). Both therapies increased the numbers of IL-17A-positive cells in the dermis of treated psoriatic lesions, although not significantly, and there was a large amount of variation between patients (Table SV). The expression of IL-17A mRNA was very low in controls’ and patients’ non-lesional samples, while in lesional skin the level of IL-17A mRNA was from 30- to 700-fold higher than in non-lesional samples (p = 0.047) (Fig. 1a). Both therapies down-regulated the expression of IL-17A; which fell to almost zero with the combination therapy (p = 0.110). In PBMCs, the expression of IL-17A mRNA was low and measurable in only a few samples (Fig. 1b).
Fig. 1. The effect of one week topical therapy with betamethasone monotherapy (Bm) and calcipotriol/betamethasone combination therapy (Cp/Bm) on the expression of interleukin (IL)-17/IL-23 pathway molecular markers. Total RNA was isolated from skin and peripheral blood mononuclear cell (PBMC) samples and the expression of (a, b) IL-17A, (c, d) IL-23, (e, f) IL-6, (g) antimicrobial peptide S100A7, and (h) chemokine CCL-20 were measured using quantitative real-time PCR. Control: healthy control; Non-lesional: skin biopsy from non-lesional skin of patient with psoriasis; Before treatment: skin biopsy from untreated lesional skin/PBMC sample before treatment; 1 week: skin biopsy/PBMC sample taken after one week of treatment with either betamethasone or calcipotriol/betamethasone; fold change: the normalized expression level compared with normal skin/PBMCs.
IL-23A is involved in the generation and activation of IL-17-producing effector cells (26, 27). The expression of IL-23A mRNA was higher in skin samples taken from psoriatic patients than controls (Fig. 1c) and significantly higher in lesional samples before treatment than in non-lesional samples (p = 0.002). Both treatments down-regulated the expression of IL-23 mRNA in lesional skin as well as in PBMCs (Fig. 1c, d), but the effect was more marked, although not significant, with the combination than the monotherapy. Control PBMCs expressed slightly more IL-23A than those of psoriatic patients’ before treatment (p = 0.396) (Fig. 1d).
IL-6 is a pro-inflammatory cytokine produced by several different cell types (28). It is essential in enhancing the development of Th17 cells. The expression of IL-6 mRNA varied greatly in skin samples and no consistent changes were observed (Fig. 1e). In control PBMCs the expression of IL-6 mRNA was higher than in patient samples (p = 0.040) (Fig. 1f). Combination therapy down-regulated the expression of IL-6 in PBMCs, but the change was not statistically significant (Fig. 1f). Monotherapy had no effect (Fig. 1e, f).
The IL-17A-related antimicrobial peptide S100A7 acts as a chemoattractant to amplify inflammation in psoriasis (17). Chemokine CCL-20 is a key stimulus for attracting CCR6+ T cells from blood to inflamed skin (29). The expression of both S100A7 and CCL-20 were near to zero in control and non-lesional skin samples, but were up-regulated in lesional samples, significantly so compared with non-lesional samples (S100A7: p = 0.000; CCL-20: p = 0.018) (Fig. 1g, h). Both therapies showed a consistent trend to down-regulate the expression of S100A7 in lesional skin (p = 0.003); combination was more efficient, although not significantly (p = 0.251) (Fig. 1g), and also reduced the expression of CCL-20 (p = 0.175); whereas monotherapy had no effect on CCL-20 (Fig. 1h).
Using FCM, the numbers of Th17 and Th22 cells as well as skin-homing Th17 cells were measured in PBMC samples. Before treatment, the median proportion of Th17 memory cells (CD4+, CD45RA-, CXCR3-, CCR6+ cells) was 15.2% (range 7.9–25.7%) in control samples and 12.4% (range 16.6–21.7%) in patient samples. The total numbers of circulating Th17 memory cells increased during both treatments, although not significantly. Combination therapy reduced the numbers of Th22 cells (CD4+, CD45RA-, CXCR3-, CCR6+, CCR10+ cells) and skin-homing CLA+ Th17 cells (CD4+, CD45RA-, CXCR3-, CCR6+, CLA+ cells) to a greater extent than monotherapy, but no statistically significant differences were observed (Fig. S2).
The expression of TNF-α mRNA in skin samples showed inter-individual variation, but both therapies down-regulated the expression in psoriatic lesions, with a more pronounced effect with monotherapy (p = 0.347) (Fig. 2a). In PBMCs, the highest expression of TNF-α (Fig. 2b) was seen in control samples (p = 0.065). The expression level of TNF-α in PBMCs was down-regulated by both therapies, although not significantly (p = 0.899) (Fig. 2b).
Fig. 2. The effect of one week topical therapy with betamethasone monotherapy (Bm) and calcipotriol/betamethasone combination therapy (Cp/Bm) on the expression of Th1 and Treg molecular markers. Total RNA was isolated from skin and peripheral blood mononuclear cell (PBMC) samples and the expression of (a, b) TNF-α, (c–d) T-bet, (e, f) IFN-γ, and (g, h) FoxP3 were measured using quantitative real-time PCR. Control: healthy control; Non-lesional: skin biopsy from non-lesional skin of patient with psoriasis; Before treatment: skin biopsy from untreated lesional skin/PBMC sample before treatment; 1 week: skin biopsy/PBMC sample taken after one week of treatment; fold change: the normalized expression level compared with normal skin/PBMCs.
Immunohistochemical staining revealed that CD3+ cells were significantly more numerous in lesional than in non-lesional and control samples (Table SIV). The results were similar and also statistically significant with anti-CD4 and anti-CD8 staining (Table SIV). Combination therapy reduced the numbers of CD3+, CD4+ and CD8+ cells markedly, but not significantly (Table SV, Fig. S1b–d). Monotherapy reduced the numbers of CD3+ and CD4+ cells in the dermis, but not in the epidermis, and not statistically significantly; also anti-CD8 staining showed no statistical difference (Table SV).
The expression of T-bet (a transcription factor promoting Th1 differentiation) was slightly, not significantly, higher in lesional skin samples than in control and non-lesional samples (Fig. 2c). Neither of the therapies had a strong effect on the expression of T-bet (Fig. 2c). However, the expression of the effector cytokine IFN-γ was more prominent in lesional than in control and non-lesional samples, and significant difference was seen between non-lesional and lesional samples (p = 0.003). Combination therapy down-regulated its expression markedly, but not significantly (Fig. 2e). Monotherapy had no effect on the expression of IFN-γ (Fig. 2e). In PBMCs, the highest, but not significantly so, expression of T-bet and IFN-γ (Fig. 2d, f) was seen in control samples. In PBMC samples from psoriatic patients, the expression levels of T-bet and IFN-γ were down-regulated by the combination therapy; betamethasone monotherapy had no effect and no statistically significant differences were observed (Fig. 2d, f).
The proportions of Th1 memory cells in PBMC samples (CD4+, CD45RA-, CXCR3+, CCR6-) decreased during both therapies, more so, but not significantly, with the combination therapy (median from 33.3% to 29.7%). Before treatment, the total CD4+ cell count was slightly higher (p = 0.221) in the control (median 41.3 % of lymphocytes; 27.5–56.3%) than in psoriatic samples (38.9%; 19.4–53.4%). Combination therapy increased the numbers of CD4+ cells, while monotherapy had no effect. No significant difference was observed between the 2 topical therapies. CD8+ cell counts did not differ between patients and controls (median 27.4%; 13.6–45.8% and 27.30%; 16.1–31.4% of lymphocytes, respectively), and combination therapy had a tendency to decrease the median number of CD8+ cells, whereas monotherapy increased it (data not shown).
FoxP3, a specific marker of natural Tregs, was used to analyse the numbers and activity of Tregs in the skin and PBMCs. Immunohistochemical staining with the FoxP3 antibody was negative in control and non-lesional skin samples in epidermis; lesional samples were positive (Table SIV, Fig. S1e). Significant differences were seen in the dermis: Tregs were most numerous in psoriatic lesions (Table SIV, Fig. S1e). While combination therapy decreased the number of Tregs (Table SV), monotherapy had little or no effect. The expression of FoxP3 mRNA in skin samples was higher in lesional than non-lesional and control samples, significantly so when non-lesional and lesional samples before treatment were compared (p = 0.017) (Fig. 2g). The combination therapy down-regulated its expression, although not significantly, while monotherapy had no effect (Fig. 2g).
The expression of FoxP3 in PBMCs was down-regulated during combination therapy, but not significantly (Fig. 2h); monotherapy increased its expression, but not significantly either. In FCM, Tregs were characterized by their CD4+, high CD25+, FoxP3+ phenotype. FCM analyses showed that patients had fewer Tregs (median: 2.0% of CD4+ cells; 0.5–3.3%) than controls (median 2.9%; 1.8–3.8%) (p = 0.086). Both therapies decreased the numbers of circulating Tregs, combination therapy more so than monotherapy (p = 0.401) (median from 1.98% to 1.64% of CD4+ cells, data not shown).
GC mediate their function through GR isoforms, GRα and GRβ being the most important clinically (30, 31). In our immunohistochemical analyses, nuclear staining in epidermis with the GRα antibody was significantly more prominent in lesional than non-lesional (p = 0.004) or control (p = 0.002) samples (Table SIV). Staining with the GRβ antibody showed no clear difference (Table SIV). Both treatments decreased the nuclear staining with both antibodies, but not significantly (Table SV, Fig. S3a, b). The expression of GRα mRNA in skin samples was higher in control than in patient samples and a statistically significant difference was observed between control samples and lesional samples before treatment (p = 0.010) (Fig. S3c). During treatment, the expression increased in lesional samples with both
therapies and, to a significantly greater extent, with the combination therapy than monotherapy (p = 0.036). The expression of GRβ varied greatly between individuals and no consistent changes were observed (Fig. S3e).
The expression of GRα was also higher in control than patient PBMCs (p = 0.257) (Fig. S3d). The expression decreased with both therapies, more so with combination therapy than monotherapy (p = 0.116). As in the skin samples, GRβ mRNA expression varied, and no far-reaching conclusion could be made (Fig. S3f).
Although biological therapies have revolutionized the treatment of severe psoriasis, most psoriatic patients are still treated with topical therapies (3, 9). The effect of the widely used calcipotriol/betamethasone treatment on the inflammatory pathways in psoriasis has been demonstrated mainly in in vitro studies (32, 33). This prompted us to compare the mechanism of action and clinical efficiency of this combination with that of betamethasone monotherapy in vivo. The derivative of betamethasone was different in the ointments studied (valerate vs. dipropionate), but we decided to use the marketed products to make this study comparable to a real-life setting.
Psoriasis is nowadays generally considered to be a TNF-α – IL-23 – IL-17-mediated disease. IL-23A, produced by antigen-presenting dendritic cells, is needed for the activation and survival of Th17 and Th22 cells and its presence is known to be significantly increased in psoriatic lesions (34). Vitamin D3 inhibits the differentiation of dendritic cells and the production of IL-23A (35). Vitamin D3 analogues should have a similar effect, and our study describes, to our knowledge, a previously unreported effect of calcipotriol/betamethasone therapy on the expression of IL-23A in vivo: the combination decreased the levels of IL-23 mRNA to a greater extent than betamethasone alone in psoriatic lesions. Th22 cells produce IL-22, which modulates psoriatic inflammation by acting as a proinflammatory cytokine on keratinocytes (8). Thus calcipotriol/betamethasone decreased the numbers of circulating Th22 cells more efficiently than betamethasone alone.
More IL-17A mRNA and IL-17A-positive cells were found in psoriatic lesions than in non-lesional and control samples. Both therapies, particularly the combination therapy, decreased the expression of IL-17A mRNA in the skin. Surprisingly, the median anti-IL-17A immunopositive cell count was elevated in treated psoriatic lesions, but the analyses were hindered by inter-sample variation, meaning firm conclusions cannot be drawn in this respect. Simultaneously the numbers of circulating skin-homing CLA+ Th17 cells was decreased, and we speculate that some of these circulating cells may have migrated to the skin during the one-week therapy. This might be explained, at least in the case of the combination therapy, by the fact that vitamin D3 enhances the ability of T cells to home to the skin by increasing numbers of CCR10 in T cells (35). It is likely that a longer period of topical therapy would have resulted in a clear decrease in the numbers of IL-17A-positive cells in psoriatic skin. The antimicrobial peptide S100A7 and the IL-17A induced chemoattractant CCL-20 are known to be overexpressed in cultured psoriatic keratinocytes (36–38); our data confirms this in vivo. IL-17A is the principal inducer of S100A7, and calcipotriol has been shown to down-regulate S100A7 expression (17). Our novel finding is that calcipotriol/betamethasone combination therapy decreased the expression of both S100A7 and CCL-20. In contrast, betamethasone monotherapy had no effect on CCL-20, and a lower impact on S100A7 than the combination. Taken together, our findings indicate that topical calcipotriol/betamethasone combination therapy down-regulates the IL-17 pathway more efficiently than betamethasone monotherapy.
Before the discovery of the IL-17A pathway, it was assumed that psoriasis was merely Th1 cell-mediated disease with an increase in the production of TNF-α and IFN-γ. TNF-α is a pro-inflammatory cytokine produced by many cell types, including keratinocytes, different dendritic cells and dermal macrophages (39). It has multiple effects in psoriasis: it regulates immune cells and the differentiation and proliferation of keratinocytes (39). In our study, both therapies decreased the expression of TNF-α in skin and PBMCs. Blocking of TNF-α has proven to be a great therapeutic option in psoriasis (6), and both therapies seem to achieve that effect. T-bet is a transcription factor that promotes Th1 cell differentiation and IFN-γ expression; the expression of T-bet correlates with the expression of IFN-γ in Th1 cells (40). In our study, both T-bet and IFN-γ mRNA levels were down-regulated in PBMCs by the combination therapy only. In the skin samples, the expression of T-bet mRNA was not altered by either therapy, but the expression of IFN-γ was down-regulated by the combination therapy. Thus, the combination therapy appears to modulate Th1 cell mediated inflammation more than betamethasone monotherapy.
Tregs have the capacity to suppress the function of different cell types and thus affect inflammatory responses. Increased presence of Tregs in psoriatic lesions compared with that in healthy skin has been described previously (41); our analysis showed a similar difference. Previously Keijsers and co-workers (42) have demonstrated that topical calcipotriol/betamethasone treatment for 8 weeks decreased the number of Tregs in psoriatic lesions and the expression of FoxP3 in the skin and PBMCs. Our findings show that treatment of only 1 week is sufficient to decrease the number of Tregs and the expression of FoxP3. In contrast, the effect of betamethasone mono-therapy on Tregs was minimal.
So far, the only published data concerning the expression of GRs in psoriasis have been from a single murine model (43) and a human study by Man and co-workers, in which GRα and GRβ were not analysed separately (44). Our analysis revealed that both isoforms are expressed in the lesional skin of psoriatic patients, with GRα nuclear immunopositivity most prominent in psoriatic lesions. Both therapies increased the expression of GRα mRNA in lesional skin, but the number of GRα immunopositive cells in skin samples and the GRα mRNA level in PBMCs were decreased. Taken together with the outcomes of our previous research (21, 45), these findings suggest that the regulation of GRα expression may be a more complex process than previously thought. Unfortunately, the current study did not increase our understanding of the function, regulation and proposed inhibitory action of GRβ (31), since its expression varied greatly in both skin and PBMCs. Therefore, in contrast with the results from the study by Vazquez-Tello et al. (19), we were unsuccessful in revealing a connection between IL-17 and IL-23 cytokines and GRβ upregulation.
In conclusion, the superior clinical efficacy of the calcipotriol/betamethasone approach could reflect a more advantageous effect on the inflammatory TNF-α – IL-23 – IL-17 axis in vivo than is provided by betamethasone monotherapy. The question remains as to whether all the changes observed were a pure effect of the topical therapies or secondary immunological alterations that manifested in the treated psoriasis lesions. We should keep in mind that additional studies with larger patient cohorts are required to address the molecular mechanisms of efficiency and side-effects seen in longer treatment periods with these treatments.
The authors are grateful for the excellent technical assistance of Ms Birgitta Grekula, Ms Anja Mattila and Ms Riitta Vuento.
This work was supported by Medical Research Center Oulu Doctoral Program, Finnish Dermatological Society, Finnish Medical Foundation, Väinö and Laina Kivi Foundation.
The authors declare no conflicts of interest.


 Click to show fullsize
Click to show fullsize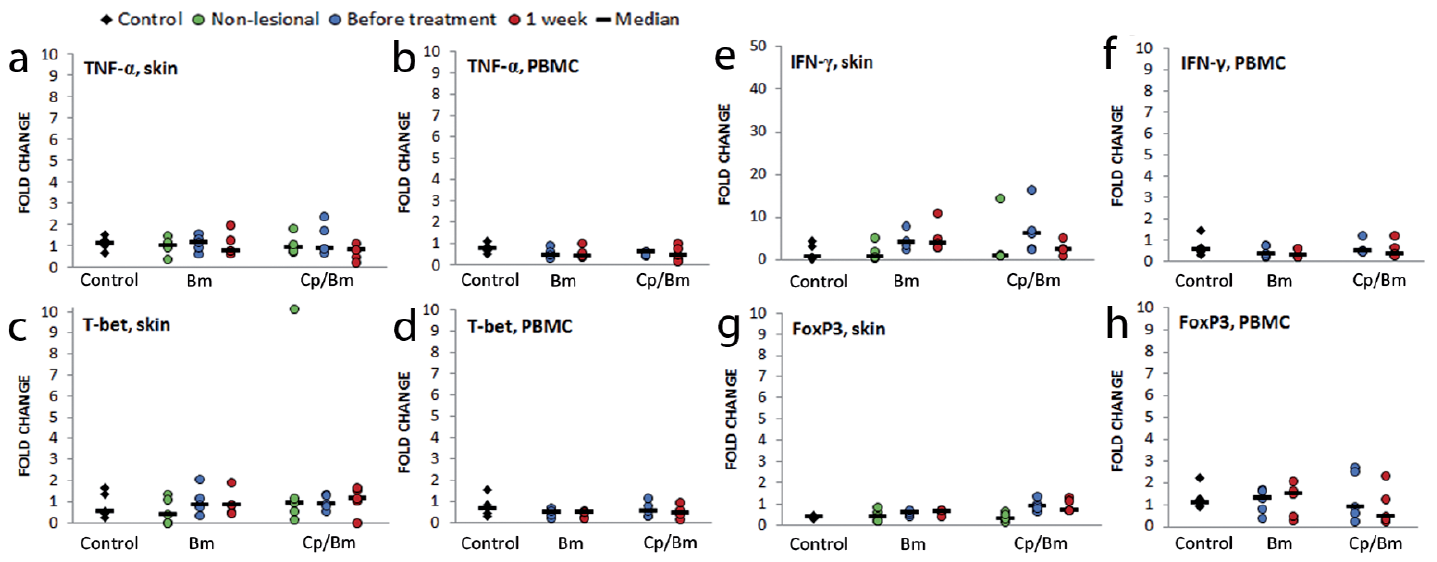 Click to show fullsize
Click to show fullsize