


Departments of 1Dermatology and Allergy and 2Clinical Immunology and Rheumatology, Hannover Medical School, Hannover, and 3University-Department of Dermatology and Allergy, Klinikum Oldenburg, Oldenburg, Germany Germany
Bullous pemphigoid (BP) is an autoimmune blistering skin disease that is more common in elderly individuals. The aim of this study was to determine the functional activity of eosinophils in patients with BP compared with healthy donors. Blood, skin and blister-derived eosinophils were strongly activated in patients with BP, seen by increased surface expression of CD69 compared with controls. CD11b was also increased in BP blood eosinophils, which may explain the striking accumulation of eosinophils in BP (1×106 per ml blister fluid). Furthermore, CCL26 was expressed by activated eosinophils in BP skin and in blister fluid. BP eosinophils also released IL-6, IL-8 and IL-1α in BP blister fluids. Apoptosis in cultivated BP eosinophils was increased and accompanied by enhanced surface externalization of CD95. Caspase 3 positive eosinophils in lesional BP skin and blister fluid also showed the initiation of apoptosis. These results reveal novel pathophysiological aspects of BP, with a strong activation pattern and increased apoptosis of eosinophils in the peripheral blood, skin and blister fluids.
Key words: apoptosis; bullous pemphigoid; CD69; eosinophils, eotaxin.
Accepted Nov 17, 2016; Epub ahead of print Nov 21, 2016
Acta Derm Venereol 2017; 97: XX–XX.
Corr: Ulrike Raap, Director University-Department of Dermatology and Allergy, Klinikum Oldenburg, Rahel-Strauss-Str. 10, DE-26133 Oldenburg, Germany. E-mail: Raap.ulrike@klinikum-oldenburg.de
Bullous pemphigoid (BP) is an autoimmune blistering skin disease that more commonly affects individuals from the age of 60 years. In general, more women than men are affected by BP (1, 2). Clinically, the early phases of BP often show flares, and sometimes additionally wheals with strong pruritus, while in late phases flares are also accompanied by tense blisters (3). The diagnosis of BP is confirmed by positive direct immunofluorescence with linear expression of C3 and IgG at the dermo-epidermal junction zone, as well as through positive indirect immunofluorescence with autoantibodies against the hemidesmosomal proteins BP180 and BP230. With regard to the extracellular membrane of BP180, a genetic predisposition has been detected in HLA-DQB1*0301 (4). Seventy percent of patients with untreated BP display elevated total IgE levels. In addition, specific IgE antibodies, reacting with the NC16A domain of BP180, have been described (5).
Due to the formation of antigen-antibody complexes a variety of different cytokines, such as IL-6 and IL-8, are released (6). Eotaxin is also secreted by epidermal keratinocytes (7), which has strong chemotactic effects on eosinophils. In patients with BP, characteristic findings are high rates of peripheral blood eosinophils, eosinophil-rich infiltrates in subepidermal blisters (8, 9), and increased numbers of tissue-resident eosinophils in skin lesions (10). This eosinophil accumulation is of particular interest, since eosinophils may contribute to the inflammation associated with BP through their release of chemokines, cytokines, and cytotoxic granule proteins. In other eosinophil-associated skin diseases, including atopic dermatitis and allergic contact eczema, the eosinophil-derived cytotoxic granule protein ECP correlates with disease severity (11–13). MCP-4, ECP, EPO and IL-5 (14, 15) have been detected in blister fluids of BP patients and support the inflammatory response. It has been demonstrated that MMP9 (16), which is released by eosinophils, plays a role in the detachment of the dermal–epidermal junction in BP (17, 18).
This study assessed the activation pattern of eosinophils in patients with BP compared with controls. Furthermore the study explored the cytokine profile of BP eosinophils and compared the apoptotic responses of BP eosinophils with those of healthy controls.
BP was defined by specific histopathological observation of subepidermal blistering, a positive indirect and direct immunofluorescence, and detection of specific auto-antibodies related to BP180 and/or BP230 (19). Patients with BP (n = 25, Table SI) did not receive systemic immunosuppressive treatment with glucocorticosteroids or diaminodiphenylsulfone or local therapy with glucocorticosteroids for the preceding 4 weeks. Blisters of the skin from patients with BP were punctured immediately after admission, analysed for eosinophil counts (Neubauer chamber) and subsequently processed to perform cytospins (1×105 cells/spot). As controls, eosinophils were obtained from the peripheral blood of 10 healthy individuals who were not atopic and had normal total IgE levels and no allergen-specific IgE-antibodies against 10 common aeroallergens. Furthermore, control blister fluids of inpatients with diseases such as bullous erysipel and phototoxic dermatitis were included (Table SII). Serum and blister fluids were centrifuged and stored at –80°C. Punch biopsies of 3-mm diameter were taken after local anaesthesia. These biopsies were embedded in Tissue-TEK® and frozen at –20°C. For subsequent studies, 5-µm thick slices were placed onto object slides.
The study was approved by the medical ethics committee of Hannover Medical School, Germany and patients gave written informed consent.
Peripheral blood eosinophils were obtained from heparin-anticoagulated venous blood and prepared following the protocol of Raap et al. (20). Briefly, blood was layered on a Ficoll density gradient (Biochrom, Berlin, Germany). After discarding the supernatant, pellets were resuspended in lysis buffer. Eosinophils were isolated from the remaining cells by a negative immunomagnetic bead selection with CD16 microbeads (Miltenyi Biotech, Bergisch Gladbach, Germany), following the manufacturer’s protocol. The purity of the eosinophils was > 98%, as assessed by fluorescence-activated cell sorting (FACS) analysis (CD66b/CD16) and Kimura staining (21). The viability was greater than 99%, as assessed by trypan blue dye exclusion. If not otherwise described, 2 × 105 eosinophils/200 µl were seeded into wells of 96 well plates and cultivated in RPMI 1640 with 10% heat inactivated foetal calf serum (FCS) including 2 mM L-glutamine, 10,000 U/ml penicillin, and 10 mg/ml streptomycin (all Seromed, Biochrom, Berlin, Germany) at 37°C and 5% CO2.
Skin sections and cytospins from blister fluids were stained following previously described protocols (22). To retrieve the antigen, slides were heated to 80°C in 0.01 M sodium citrate solution, pH 9.0, in a water bath for 30 min. Thereafter, non-specific binding sites were blocked using phosphate-buffered saline (PBS) containing 0.2% Triton X-100 (Sigma), 0.02% sodium azide (Merck, Darmstadt, Germany) and 5% normal goat or normal donkey serum for 1 h at room temperature (RT). Incubation with primary antibodies against targets, diluted in PBS containing 0.5% lambda-carrageenan (Sigma) and 0.02% sodium azide, was carried out at 4°C overnight. After washing in PBS, appropriate secondary antibodies (Jackson ImmunoResearch Europe, Suffolk, UK) diluted 1:200 in PBS-carrageenan solution were applied for 2 h at RT. After subsequent washing in PBS, cell nuclei were stained for 10 min at RT with DAPI (Sigma) and sections were mounted in anti-quenching medium. To test the specificity of the staining, primary antibodies were replaced by an equivalent amount of non-immune IgG derived from the same species as the specific antibody (negative control).
The following antibodies were used: goat polyclonal anti-mouse eotaxin 3/CCL26 antibody (ab83505, Abcam), rabbit polyclonal anti-human caspase 3 antibody (ab13847, Abcam), rabbit polyclonal anti-human major basic protein antibody (MBP, ab187523, Abcam), Mouse monoclonal anti-human CD69 antibody (Clone: FN50, ab51862, Abcam), the respective isotype controls and secondary antibodies. Nuclei staining was performed with DAPI (Abcam). Analysis was performed using an immunofluorescence microscope (Axiolab, Carl Zeiss, Jena, Germany).
Isolated eosinophils were incubated at 4°C with 100 µl blocking buffer (heat inactivated human IgG, gelatin in PBS) for 15 min. Surface antibodies were added, using FITC-conjugated mouse anti-human monoclonal CD95 (clone: UB2, Beckmann Coulter, Krefeld, Germany), PE-conjugated mouse anti-human monoclonal CD69 (clone: L78, BD Bioscience, San Jose, CA, USA), FITC-conjugated anti-human monoclonal CD11b (clone: Bear1, Abcam, Cambridge, UK) and respective IgG1-FITC and IgG2b-PE isotype controls (Beckmann Coulter) in a concentration of 2 ng/ml each. After incubation for 1 h at 4°C, eosinophils were analysed with FACSCalibur (BD Bioscience).
Briefly, eosinophils were incubated with Annexin-V and propidium iodide at 4°C for 15 min (BD Bioscience Pharmingen) and analysed by flow cytometry.
Using commercially available enzyme-linked immunoassay (ELISA) kits IL-1α, IL-6, IL-8, ECP, CCL11 and CCL26 levels were analysed in blister fluids, serum, and in supernatants from cultivated eosinophils of patients with BP according to the manufacturer’s instructions (R&D Systems, Abingdon, UK: CCL26, CCL11, IL-1α, IL-8; eBioscience Inc., Headquarters, San Diego, CA, USA: IL-6, Thermo Fisher Scientific/ Phadia, Freiburg Germany: ImmunoCAP ECP). CD95-ligand levels were analysed in blister fluids and in sera (Abcam). The analysis was performed by the FLUOstar OPTIMA Microplate Reader (BMG Labtech GmbH, Ortenberg, Germany). Detection levels were as follows: IL-6: 3.13–200 pg/ml; IL-1α, CD95-ligand 1.37–1,000 pg/ml; CCL11: 15.6–1,000 pg/ml; IL-8: 31.25–2,000 pg/ml; CCL26: 62.5–4,000 pg/ml, ECP: 2–200 µg/l.
All data are presented as means ± standard errors of the mean (SEM). Statistical analysis was performed with GraphPad Prism 5. Data were tested for their statistical distribution. Normally distributed data were analysed either with a Student’s t-test or a one-way analysis of variance (ANOVA) followed by a Tukey post-test. Non-parametric tests were used for non-normal data. For 2 groups the Wilcoxon rank-sum test or Mann-Whitney test were used, and for more than 2 groups of data the Kruskal–Wallis test followed by Dunn’s post-test was employed. Two-way ANOVA was used followed by a Bonferroni post-test. A p-value < 0.05 was considered statistically significant.
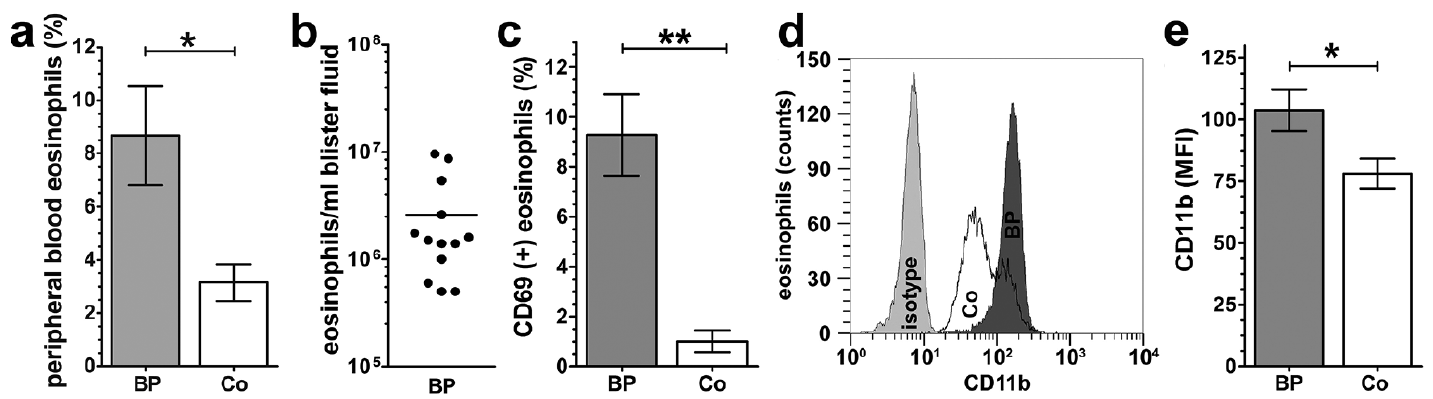
Eosinophilia has been described as a characteristic of BP (9). The current study confirmed a significant blood eosinophilia in patients with BP in comparison with healthy donors (Fig. 1a, p < 0.05). Furthermore, this study showed a strong accumulation of eosinophils in blister fluids of BP patients. The blister aspirates contained a striking number of 2.6 ± 0.8 × 106 eosinophils per ml blister fluid in patients with BP (Fig. 1b).
Fig. 1. Characterization of eosinophils from patients with bullous pemphigoid (BP). (a) Human peripheral blood eosinophils from patients with BP (BP) and healthy donors (Co) were counted (BP: n = 18; Co: n = 21; standard error of the mean (SEM); Mann–Whitney test, *p < 0.05). (b) Eosinophils in blister fluids of patients with untreated BP were counted and displayed as eosinophils number/ ml blister fluid (n = 13). (c) Fluorescence-activated cell sorting (FACS) analysis of CD69-positive eosinophils from patients with BP and healthy donors, isolated from peripheral blood and stained as described in the methods (Mann–Whitney test, BP: n = 7; Co: n = 9; SEM; **p < 0.001) (d–e) Peripheral blood eosinophils from BP and Co were stained with antibodies against CD11b and analysed by FACS. (d) Surface staining of CD11b on freshly isolated eosinophils displayed as a histogram (1 representative out of 6). (e) Mean fluorescence intensity (MFI) of CD11b expression on eosinophils (Student’s test, n = 7; SEM; *p < 0.05).
Activation levels of eosinophils from patients with BP were tested by analysing the expression of CD69, which is a well-established marker for eosinophil activation (23). Here, we observed significantly increased expression of CD69 in peripheral blood eosinophils of BP patients compared with healthy controls (Fig. 1c, p < 0.01).
Furthermore, we investigated the activation pattern of eosinophils in lesional skin and blister fluids using immunofluorescence. To identify eosinophils in the lesional skin and in cytospins, eosinophils were stained with antibodies against MBP, which is a specific marker for eosinophils (24). Distinct expression of CD69 was observed on eosinophils from lesional skin (Fig. 2a, b, d) and in blister fluids of patients with BP (Fig. 2e, g), whereas isotype control staining was negative (data not shown). In contrast, blister fluids of control patients revealed no eosinophil accumulation and no CD69 positivity (data not shown).
Fig. 2. CD69-positive eosinophils in perilesional skin and blister fluid of patients with bullous pemphigoid (BP). Immunofluorescence of perilesional skin and blister fluid-derived eosinophils from 1 representative patient with BP out of 4 different donors. Sections were stained for CD69 (red) and major basic protein (MBP) (green). Nuclei staining was performed with DAPI. (a–d) Immunofluorescence of perilesional skin. (e–g) Immunofluorescence of eosinophils from blister fluid (magnification: (a) 200×; (b–g) 400×).
Since eosinophils express at least one of the eotaxins in skin diseases (25) and given that the predominant immune response in BP is of Th2 type (26), we analysed the expression of CCL26 in CD69(+)/ MBP(+) eosinophils. We found CCL26 expression in activated eosinophils in BP lesional skin (Fig. 3a) as well as in the respective blister fluid (Fig. 3b).
Fig. 3. Major basic protein (MBP)-positive eosinophils in perilesional skin and blister fluid express CCL26. Immunofluorescence (IF) of perilesional skin and blister fluid-derived eosinophils from the same patient with bullous pemphigoid in Fig. 2. Sections were stained for CCL26 (yellow) and MBP (green). Nuclei staining was performed with DAPI (blue). IF staining of (a) perilesional skin and (b) eosinophils from blister fluid (magnification: 400×).
The expression levels of CD11b, a factor involved in the adhesion and transmigration of eosinophils (27), were tested to determine whether they were enhanced in eosinophils from BP lesional skin and blister fluids. Surface expression of CD11b was higher on BP eosinophils in comparison with eosinophils from healthy donors (Fig. 1d–e, p < 0.05). A representative histogram from the FACS analysis is shown in Fig. 1d.
As shown above, we demonstrated that BP eosinophils are highly activated in the peripheral blood, skin and blister fluid of patients with BP and tissue-derived CD69-positive eosinophils express CCL26. Various chemokines and cytokines are elevated in blister fluid and sera of patients with BP (14). Therefore, we tested the levels of cytokines and chemokines in blister fluid and sera in comparison with those in the supernatant of cultivated peripheral blood eosinophils from patients with BP. We first cultured 1×106 peripheral blood eosinophils per ml from patients with BP for 24 h and then analysed the supernatant for cytokine content by ELISA in parallel with cytokine levels in respective blister and serum fluids. Here, elevated levels of IL-6 (p < 0.001), IL-8 (p < 0.001), IL-1α (p < 0.05), CCL11 (p < 0.001) and ECP (p < 0.05) were detected in the blister fluids compared with sera (Fig. 4a–e). Similar observations were also made regarding CCL26 (Fig. 4f). Comparing blister fluids with supernatants, we found significantly higher levels of IL-6 (p < 0.01), IL-8 (p < 0.01) and CCL11 (p < 0.001) in blister fluids (Fig. 4a, b, e), whereas ECP levels were higher in supernatants compared with blister fluids (p < 0.01, Fig. 4d). Comparing sera with supernatants, increased levels of IL-6, IL-1α and ECP were also found in supernatants, although this increase was not statistically significant (Fig. 4a, c, d,). IL-8 was not detectable in sera. CCL11 and CCL26 levels were both higher in sera than in supernatants (Fig. 4e, f, p < 0.05).
Fig. 4. Analysis of interleukins and chemokines in blister fluids, serum and supernatant of eosinophils derived from patients with bullous pemphigoid. Blister fluids, serum samples and supernatants of unstimulated and isolated peripheral blood eosinophils (2 × 105, 24 h) from untreated patients with BP (n = 6–15) were analysed for IL-6 (a), IL-8 (b), IL-1α (c), ECP (d), CCL11 (e) and CCL26 (f) (SN: supernatants; SEM; 1-way analysis of variance (ANOVA): F(2/15)=25.33, p < 0.0001 (IL-6), F(2/19) = 15.50, p = 0.0001 (IL-8), F(2/16) = 20.33, p < 0.0001 (CCL11), Tukey post hoc, Kruskal–Wallis test: p = 0.0151 (IL-1α), p = 0.0014 (ECP), p = 0.0129 (CCL26), Dunn post hoc, *p < 0.05; **p < 0.01; ***p < 0.001).
To determine whether the increased number of eosinophils in the peripheral blood of patients with BP display delayed apoptosis, similar to eosinophils from patients with atopic dermatitis (28), we determined the apoptosis of eosinophils from patients with BP in comparison with healthy controls. After 24 h of culture we observed a distinct increase in apoptotic eosinophils in patients with BP compared with healthy controls (Fig. 5b, c, p < 0.01), while no significant difference between eosinophils from patients with BP and controls was seen directly after isolation (Fig. 5a, c).
Fig. 5. Increased apoptosis in eosinophils from patients with bullous pemphigoid. Analysis of apoptosis of peripheral blood eosinophils from BP patients (BP) and healthy donors (Co) after isolation (a, 0 h) and cultivation (b, 24 h). One FACS analysis representative for seven individual experiments is displayed. c) Combined results of the FACS analysis illustrating the mean percent of apoptotic eosinophils; BP: n = 7; Co: n = 8; SEM; *p < 0.05, **p < 0.01, ***p < 0.001). d) Analysis of CD95 displayed as MFI after isolation (0 h) and 24 h culture (n = 6; SEM; *p < 0.05). e) Analysis of CD95 expression regarding the percentage of positive eosinophils after isolation (0 h) and 24 h culture (n = 6; SEM; *p < 0.01). f) Measurement of the CD95 ligand in serum and blister fluids from BP patients (n = 14; SEM; **p < 0.01).
Since the CD95 surface receptor and CD95 ligand are members of the signal cascades leading to apoptosis, we investigated the expression of CD95 on peripheral blood eosinophils from patients with BP. We did not find any difference in the expression of CD95 on eosinophils from patients with BP compared with healthy controls directly after isolation, either in terms of mean fluorescence intensity (MFI, Fig. 5d) or in the percentage of CD95(+) eosinophils (Fig. 5e). However, we did observe significantly higher surface expression of CD95 on blood eosinophils from patients with BP compared with healthy controls after 24 h of culture based on the MFI (Fig. 5d, p < 0.05) as well as on the percentage of CD95(+) eosinophils (Fig. 5e). Because of these observations, we measured the concentration of CD95-ligand in blister fluids and sera of patients with BP and found that levels of this ligand were significantly higher in blister fluids in comparison with sera of patients with BP (Fig. 5f, p < 0.01). In contrast, CD95-ligand was not detectable in either control blister fluids or sera (data not shown). Because in vitro culture of eosinophils showed increased apoptosis we next analysed the state of apoptosis of eosinophils in BP skin and blister fluid. The expression of caspase 3 is an early sign of apoptosis (29). Therefore, skin biopsies and cytospins of blister fluids from patients with BP were stained with antibodies against MBP and caspase 3. Tissue-resident and blister fluid-derived MBP positive eosinophils were clearly positive for caspase 3 (Fig. 6a–f).
Fig. 6. Caspase-3-positive eosinophils in perilesional skin (a–c) and blister fluid (d–f). Immunofluorescence of perilesional skin and blister fluid-derived eosinophils from 1 representative BP patient out of 4 different donors. Sections were stained for Caspase 3 (orange) and MBP (green). Nuclei staining was performed with DAPI (magnification: (a–c) 200×; (d–f) 400×).
This study shows, for the first time, that eosinophils in sera, blister fluids and the skin of patients with BP are highly activated, as demonstrated by the expression of the activation marker CD69. The study also shows that eosinophils accumulate strongly in blister fluids and that their numbers are increased in the peripheral blood accompanied by elevated CD11b expressions. In addition, tissue and blister eosinophils are positive for eotaxin. Further, increased levels of IL-6, IL-8 and IL-1α were found in blister fluids of patients with BP. Interestingly, although patients with BP show a strong accumulation of eosinophils in the peripheral blood, skin and blister fluid, these eosinophils display early signs of apoptosis in the skin and blister fluid, as seen by increased caspase 3 expression, as confirmed by externalization of CD95 and increased expression of Annexin V in peripheral blood eosinophils.
In our study we show that approximately 3 million eosinophils accumulate per ml blister fluid in patients with BP. Previously, increased eosinophils have been described only by histology but not by counting eosinophils in blister fluids directly (8). This extensive accumulation of eosinophils has great pathophysiological impact on the local skin inflammation in patients with BP and might be a key causative factor for localized itch. Given the accumulation of eosinophils at lesional sites it is interesting that strongly activated peripheral blood eosinophils of patients with BP show increased expression of CD11b, which is involved in the transmigration and adhesion of eosinophils (27).
The novelty of the current study is that, in comparison with blister fluids and sera, we also analysed the supernatants of cultured eosinophils derived from the same donor as the blister fluid and sera for cytokine levels, including IL-6, IL-8, CCL11, and 26, ECP and IL-1α of each patient with BP. We confirmed data by other groups (30, 31) showing high levels of IL-6 and IL-8 in blister fluid and sera of patients with BP. The increased levels of IL-6 and IL-8 in the supernatants of cultivated BP eosinophils show that eosinophils, in addition to keratinocytes, may have a role in the production of these cytokines in the blister fluids. So far, IL-6 is known to influence the Th1/Th2 differentiation (32) as well as the induction of histamine production (33) and IL-8 is a chemoattractant for neutrophils (34). The expression of these cytokines is interesting because the development of bullous lesions is dependent on the cutaneous infiltration of activated autoreactive CD4+ T cells, eosinophils and neutrophils (4, 35–37).
Our observations regarding increased levels of CCL11 and CCL26 in blister fluids and sera of patients with BP are consistent with other reports (7, 38). The expression of the eosinophil chemoattractants CCL11 by peripheral blood eosinophils and CCL26 by tissue-resident eosinophils seen in our study may explain not only the enhanced accumulation of eosinophils, but also of CD4+ T cells and neutrophils, in the lesional skin. Since CCL26 blocks the recruitment of Th1 lymphocytes via CCR1 and CCR5, the expression of CCL26 by eosinophils may augment the Th2-immune response in BP (39).
ECP is involved in pore formation in target cell membranes (40). Thus, high concentrations of ECP in BP blister fluids and in the BP eosinophil supernatants, as shown in our study, provide further evidence for a pivotal role of eosinophils in BP.
In addition, we found low levels of IL-1α in BP blister fluids, which is in accordance with previously published data (30). Although the concentration in blister fluid was low, the level of IL-1α expressed by isolated peripheral eosinophils was approximately the same as in the blister fluids. This finding is may be highly relevant in terms of the severity of inflammation and blister formation in patients with BP, given the potent pro-inflammatory actions of IL-1α, (41). IL-1 α activates gelatinase B (MMP9) (42), which is a critical component of blister formation in patients with BP (43, 44). Gelatinase B is expressed by neutrophils, but also eosinophils (44). Recently, the expression of gelatinase B by eosinophils was described in patients with squamous cell carcinoma, whereas eosinophils from healthy subjects express very low levels of this enzyme (45–47). Since migration to the basement membrane zone in an IgG-dependent manner without split formation was shown for the eosinophil cell line 15HL-60, it would be interesting to analyse gelatinase B expression as a possible causative factor for split formation in eosinophils from patients with BP.
Besides the activation of eosinophils and their cytokine profile, we showed that the number of peripheral blood eosinophils from patients with BP is substantially increased, but they also display higher levels of apoptosis than healthy donor eosinophils. Furthermore, BP peripheral blood eosinophils show an increased surface expression of the cell death receptor CD95 in comparison with healthy donor eosinophils. This finding is interesting, given the fact that in atopic diseases, such as atopic dermatitis, the peripheral blood eosinophilia is associated with an inhibition of apoptosis (48, 49). This led us to hypothesize that CD95 ligand-mediated apoptosis may play a role in the regulation of eosinophil survival in patients with BP. The activation of the CD95/CD95L signalling cascade is specifically activated by the membrane-bound CD95-ligand (mCD95), which is expressed on cells such as T cells, neutrophils and monocytes (50, 51), but also by the soluble CD95-ligand (sCD95) (50, 52). The concentration of sCD95-ligand in the sera from patients with BP was elevated compared with control patients, providing an explanation for the increased apoptosis of peripheral blood eosinophils in patients with BP compared with controls. The enhanced apoptosis was further confirmed by the expression of caspase 3 in tissue-resident and blister fluid-derived eosinophils of patients with BP.
Eosinophils in sera, skin and blister fluids from patients with BP are highly activated and display increased apoptosis in both blister fluids and skin. Eosinophil accumulation in skin and blisters may be explained by an increased expression of CD11b on peripheral blood eosinophils. Furthermore, BP eosinophils are able to secrete inflammatory cytokines and granule proteins, which, in turn, modulate the immune reaction in BP. It is notable that the accumulation of eosinophils in BP is not associated with an inhibition of apoptosis, as displayed in other inflammatory skin diseases, such as atopic dermatitis. For upcoming studies, the NC14A knockout mouse model (53), which develops a similar BP phenotype, may be employed to evaluate the modulation of the immune reaction by eosinophils in more detail. Together, the data from this study add novel aspects regarding the functional activity and role of eosinophils in patients with BP.
This work was supported by a DFG grant to Ulrike Raap RA 1026/1-2 as part of the KFO250. Judith Engmann was supported through funds of the Hannover Biomedical Research School DFG, GSC 108. The authors would like to thank S. W. Evers, (University of Oxford) and Professor B. F. Gibbs (University of Kent) for English proofreading of the manuscript.


 Click to show fullsize
Click to show fullsize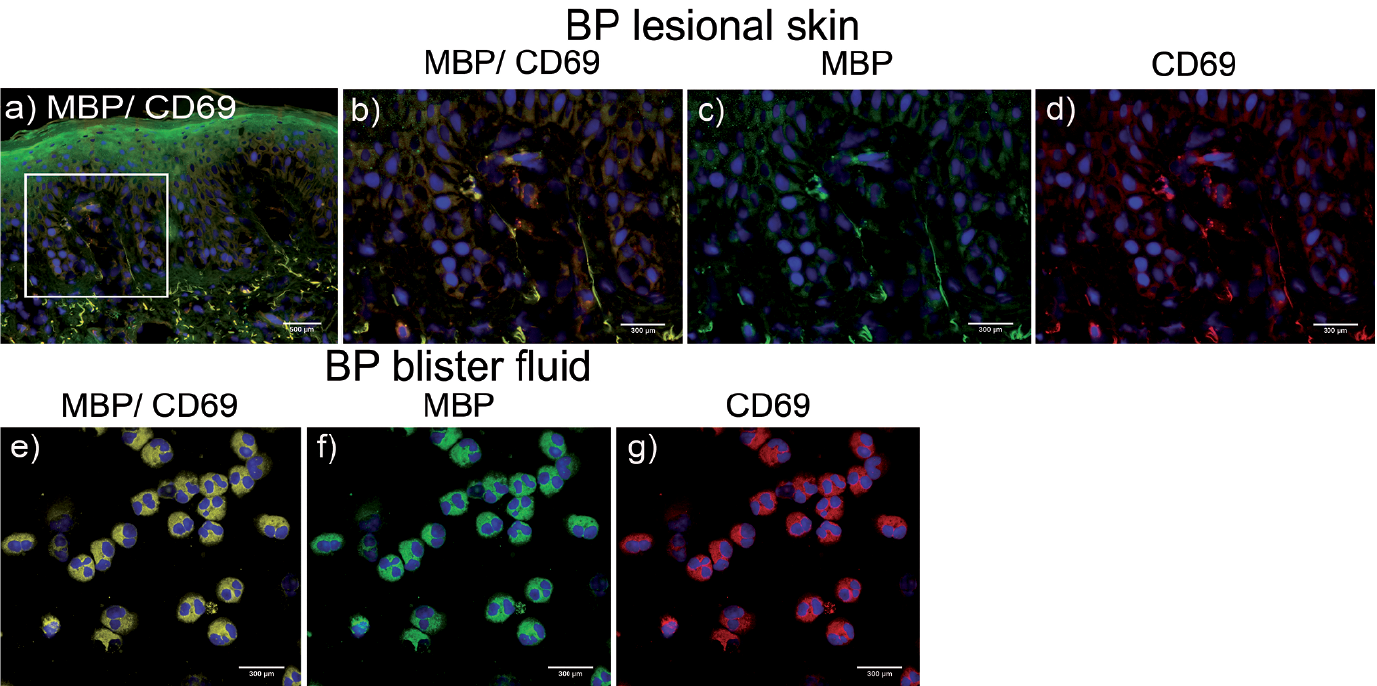 Click to show fullsize
Click to show fullsize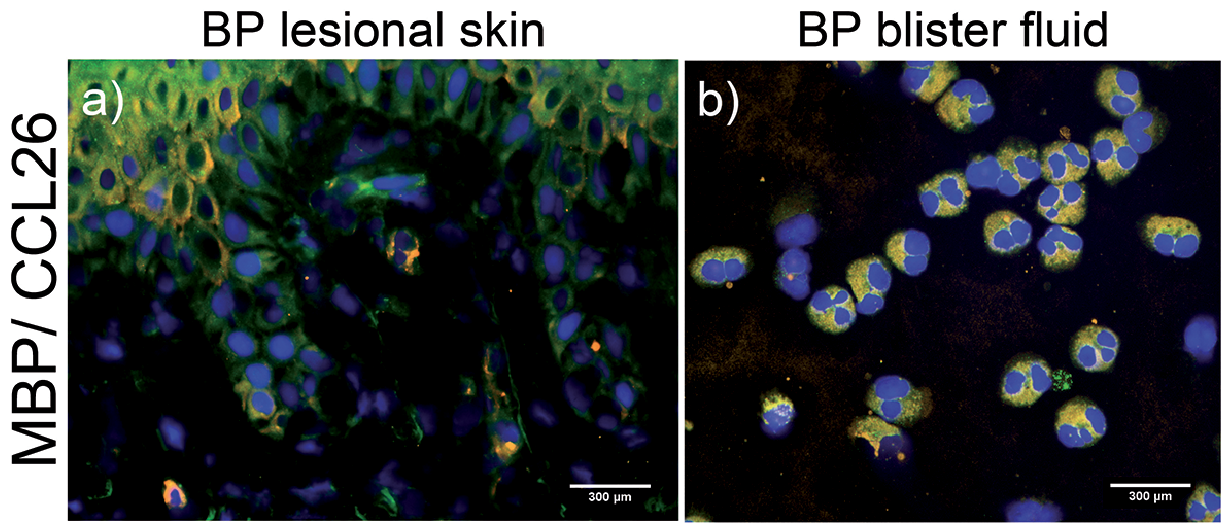 Click to show fullsize
Click to show fullsize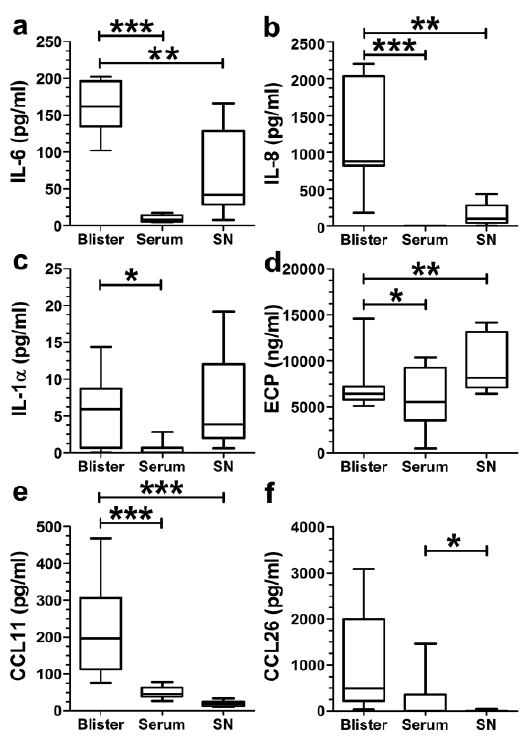 Click to show fullsize
Click to show fullsize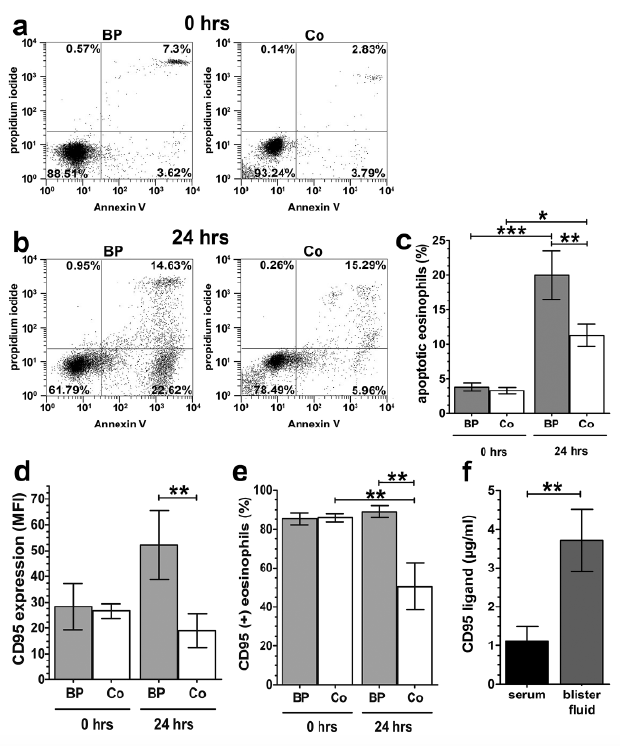 Click to show fullsize
Click to show fullsize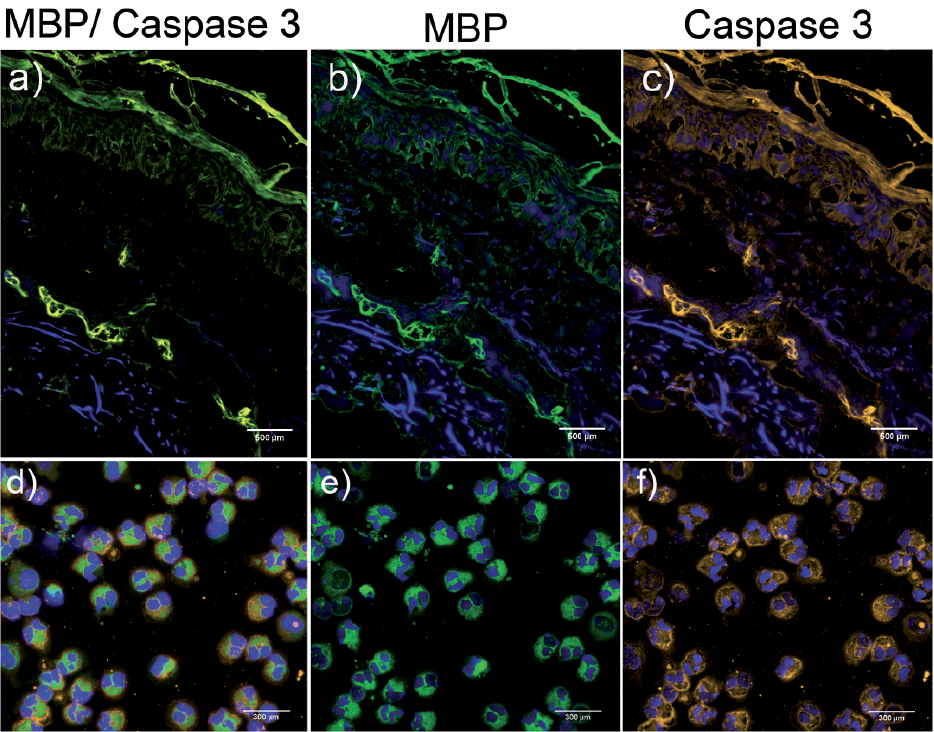 Click to show fullsize
Click to show fullsize