


1Laboratory of Immunogenetic and Tissues Immunology, Department of Clinical Immunology, 5Laboratory of Tumor Molecular Immunobiology, Department of Experimental Oncology and 8Laboratory of Tissues Immunology, Ludwik Hirszfeld Institute of Immunology and Experimental Therapy, Polish Academy of Sciences, 2Department of Dermatology, Venerology and Allergology, Silesian Piast Wroclaw Medical University, Wroclaw, 3Department of Biochemistry and Molecular Biology, Karol Marcinkowski Medical University of Poznan, Poznan, 4Department of Histology, Medical College, Jagiellonian University, Cracow, Poland, 6School of Biomedicine, Far Eastern Medical Center, Far Eastern Federal University, Vladivostok, Russia, and 7Department of Immunology, MD Anderson Cancer Center, The University of Texas, Houston, TX, USA
Psoriasis is an inflammatory immunogenetic skin disease, often accompanied by itch. Opioid receptors are known regulators of itch sensation in the central nervous system. In the brain, μ-opioid receptors may potentiate itch, while activation of κ-opioid receptors may reduce or even alleviate itch; however, the role of opioid receptors in itch perception in the skin is
poorly understood. To further elucidate the role of opioid receptors in the neurobiology of psoriatic itch, punch biopsies of non-lesional and lesional skin of
patients with psoriasis and healthy controls were
studied. Real-time polymerase chain reaction and immunofluorescence microscopy were used to detect opioid receptor genes and protein expression, respectively. The OPRK1/κ-opioid receptor pathway was found to be downregulated in lesional skin of psoriasis, correlating positively with itch sensation. In contrast, the OPRM1/μ-opioid receptor system was uniformly expressed by epidermal keratinocytes in all analysed groups. These findings suggest that imbalance of epidermal opioid receptors may result in disordered neuroepidermal homeostasis in psoriasis, which could potentiate transmission of itch.
Key words: itch; psoriasis; opioid receptor; keratinocyte, neuro-inflammation; neuropeptide.
Accepted Dec 12, 2016; Epub ahead of print Dec 13, 2016
Acta Derm Venereol 2017; 97: XX–XX.
Corr: Jacek C. Szepietowski, Department of Dermatology, Venerology and Allergology, Silesian Piast Wroclaw Medical University, PL-50-368 Wroc?aw, Poland. E-mail: jacek.szepietowski@umed.wroc.pl
Psoriasis is a multisystem inflammatory skin disease with an immunogenetic background and a prevalence of 1–3% in the general population (1, 2). Although the exact pathomechanism of psoriasis is not fully understood, it is suggested that communication between the skin, endocrine, and immune systems creates the molecular basis for this condition (1, 3). Psoriatic epidermal keratinocytes and immune cells may secrete a wide spectrum of neuropeptides, which modify adaptive and innate mechanisms of immune response. Skin lesions in patients with psoriasis are often accompanied by itch; approximately 70–90% of patients with psoriasis have this unpleasant sensation (4). The molecular basis for psoriatic itch is unknown; however, the disordered interplay between skin, immune cells, genetic and neurological systems with decreased psychoneurophysiological tolerance is considered to be the main factor potentiating itch transmission (4–7). Recent data suggest that the endogenous opioid system (EOS) and peripheral innervations of axonal nerve terminals of neurones from dorsal root ganglions (DRG) may participate in itch transmission (8). The following opioid receptors (ORs; gene/protein) have been identified: OPRM1/µ-opioid receptor (MOR), OPRK1/κ-opioid receptor (KOR), OPRD1/delta-opioid receptor (DOR) and OPRL1/opioid growth factor receptor-like 1 (OGFRL1), which belong to the G-protein-coupled receptors (GPCR), rhodopsin subtype family (9, 10). Central nervous system (CNS) neurobiological studies using functional magnetic resonance imaging (fMRI) have shown that both MOR and KOR systems are involved in central itch regulation (11–15). However, it is not known whether the peripheral EOS, expressed by skin components and nerve terminals, participates in initiating and transmitting itch signalling between the skin and the brain (3, 16, 17).
Based on earlier experimental and clinical observations it is suggested that the OPRM1/MOR system supports itch transmission, and skin neuroinflammation promotes its expression. In contrast, the OPRK1/KOR system probably operates in opposition to OPRM1/MOR and inhibits itch transmission in the skin, although neuroinflammation may destabilize its expression (18–21).
The aim of the current study was to further assess gene and protein expression of ORs in skin biopsies of patients with psoriasis with and without itch.
Skin tissue punch biopsies were collected from 20 psoriasis inpatients (4 females and 16 males; age range 24–65 years; mean age 44.1 ± 12.5 years). Exclusion criteria were: patients who had received any local or general systemic antipsoriatic therapy within 3 months prior to entering the study and/or had any other cutaneous or systemic disease that might influence the results. The control group comprised 20 healthy subjects (12 females and 8 males) who had undergone routine scheduled plastic surgery in the Plastic Surgery Unit of the Department of Dermatology, Venereology and Allergology, Silesian Piast Wroclaw Medical University. The age range of the control group was 30–79 years (mean age 56.3 ± 13.0 years).
All patients provided written informed consent to participate in the study. The study was carried out accordance with good clinical and laboratory practices and the principles of the Declaration of Helsinki, and was approved by the local ethics committee of Wroclaw Medical University (669/2011).
All patients were evaluated for clinical parameters. Psoriasis severity was evaluated using the Psoriasis Area Severity Index (PASI) and itch intensity was assessed with a visual analogue scale (VAS). All procedures were performed in accordance with the recommendations of the Psychodermatology Section of the Polish Dermatological Society. Punch biopsies were taken from non-lesional and lesional skin for analysis of OR gene and protein expression. Prior to gene analysis, the skin biopsies underwent homogenization, RNA isolation, and reverse transcription on cDNA. Multiplex real-time PCR (RT-PCR) for OPRM1 and OPRK1 genes was applied, with ACTB as housekeeping gene, and relative gene expression analysis was carried out. MOR and KOR opioid receptors were detected in frozen skin sections using an indirect immunofluorescence protocol with primary rabbit polyclonal antibodies recognized ORs and secondary conjugated fluorochrome antibodies. OR expression was evaluated using image semiquantitative analysis, based on immunofluorescence intensity. All data are presented as medians with lower and upper quartiles. A detailed description of the Materials and Methods is given in Appendix S1.
The mean psoriasis area and severity index (PASI) in the studied patients was 15.6 ± 9.9 points. Fourteen patients reported itch (mean VAS 3.9 ± 2.7 points) (Itch+), while the remaining 6 did not have itch (Itch–).
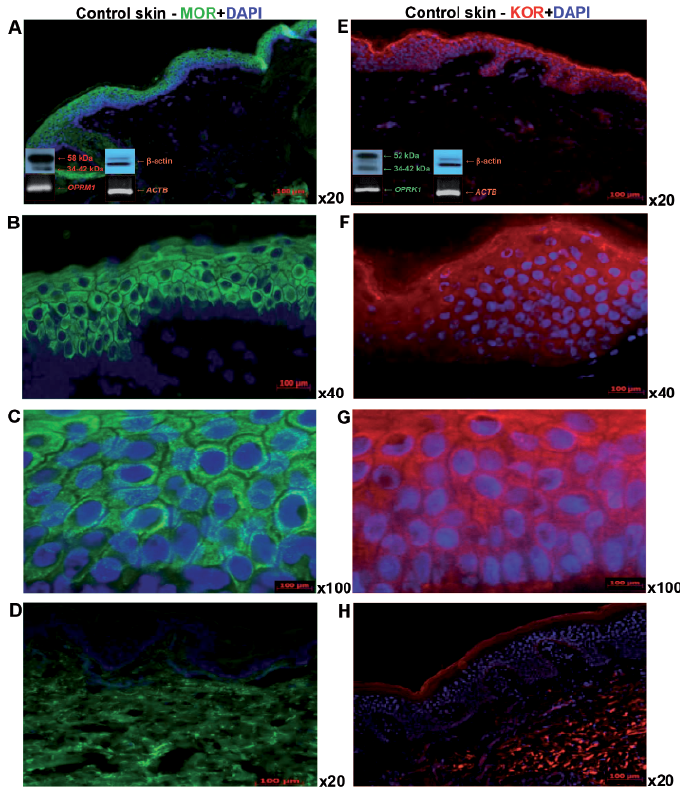
All fresh-frozen skin samples from the control group had detectable protein levels of MOR (Fig. 1A–C) and KOR (Fig. 1E–G) , with the same pattern of cytoplasmic and membrane staining. MOR was expressed predominantly in the upper layers of the epidermis, and at a lower level in the stratum basale. In the dermis, MOR expression was absent, with minor exceptions for single fibroblast-like cells, mononuclear cells and nerve fibre structures (Fig. 1A–C). The OPRM1 gene transcript in the skin was expressed at a very low level; practically at the limit of RT-PCR sensitivity, but its specific expression was detectable in all skin samples analysed (Fig. 1A).
Fig. 1. Expression of μ-opioid receptor (MOR) and κ-opioid receptor (KOR) with DAPI counterstaining in normal skin of control group. Immunofluorescence of epidermal MOR expression in normal healthy skin of control group: magnification (A) ×20, (B) ×40 and (C) ×100. (D) Omission of MOR primary antibody, (E) immunofluorescence of epidermal KOR expression: magnification (E) ×20, (F) ×40 and (G) ×100; and (H) omission of KOR primary antibody. Western blots using the same (A) MOR and (E) KOR antibody and reference for β-actin (bottom left corner, above), showing OR protein expression, below: real-time PCR amplificated transcripts confirmed for OPRM1 (A) and OPRK1 (E) and reference gene ACTB (A, E), respectively, using 2,5% agarose gel electophoresis and ethidum bromide for bands visualization. DAPI: (4’,6-diamidino-2-phenylindole)
KOR was expressed uniformly by all epidermal layers, with weak signals detectable in the dermis (Fig. 1E–H). Antibodies used in western blotting recognized external tails of MOR and KOR receptors and their nuclear and membrane forms, respectively (Fig. 1A, E). The OPRK1 gene transcript was also expressed at a low level in the skin, but, in contrast to OPRM1, its expression was higher and was also detectable in all analysed skin samples (Fig. 1E). The amplification control for both OR genes was obtained using total RNA from human brain reference material (Agilent Technologies, Santa Clara, USA) (data not shown). Omission of the primary antibodies for both OR proteins did not result in any fluorescence signals (Fig. 1H). Similarly, addition of water or a non-reverse-transcribed RNA sample prevented the detection of any amplification activity in RT-PCR (data not shown).
Not significantly different OPRM1/MOR expression was demonstrated in all analysed psoriasis samples from non-lesional and lesional skin (Figs 2a, b and 3). In comparison with 20 control group subjects, no statistically significant differences in OPRM1/MOR levels were observed (Fig. 2a, b). The OPRM1/MOR system was expressed uniformly in samples of non-lesional and lesional skin of patients with psoriasis with or without itch and in the control group (Figs 3 and 4a, b). OPRM1/MOR expression did not show any significant relationship to clinical parameters studied, such as disease severity or itch intensity, based on PASI and VAS in all skin groups (data not shown).
Fig. 2. Expression of MOR/OPRM1 (a and b, respectively) and KOR/OPRK1 (c and d, respectively) opioid receptors in control group and non-lesional and lesional skin of patients with psoriasis (**p < 0,01; ***p < 0.001).
Fig. 3. Expression of µ-opioid receptor (MOR) with DAPI counterstaining in non-lesional and lesional skin of patients with psoriasis with itch (Itch+) and without itch (Itch–). Immunofluorescence of epidermal MOR expression in frozen skin sections from patients with psoriasis in (A, B) non-lesional and (C, D) lesional skin with itch (Itch+) (×20 and ×40 magnification, respectively), in comparison with (E, F) non-lesional and (G, H) lesional skin without itch (Itch–) (magnification ×20 and ×40, respectively). Western blotting using the same MOR antibody (bottom left corner); non-lesional skin (A) with and (C) without itch in comparison with lesional skin (E) with and (G) without itch. DAPI: 4’,6-diamidino-2-phenylindole.
Fig. 4. Expression of MOR/OPRM1 (a and b, respectively) and KOR/OPRK1 (c and d, respectively) opioid receptors in the skin of control group and non-lesional and lesional skin of psoriasis with and without itch (*p < 0.05, **p < 0.01, ***p < 0.001).
For OPRK1/KOR expression, a significant decrease in OPRK1 transcript expression was observed in non-lesional and lesional skin of patients with psoriasis compared with the control group (Fig. 2d; p < 0.01). A significant decrease in KOR expression was limited to the lesional skin in patients with psoriasis and to the control group (p < 0.01; Fig. 2c). Non-lesional KOR expression was significantly increased compared with control skin (Fig. 2c; p < 0.01) and correlated positively with PASI score (r = 0.53; p = 0.01) (data not shown).
Statistically significant differences were observed in the OPRK1/KOR system in relation to itch (Fig. 5).
Fig. 5. Expression of κ-opioid receptor (KOR) with DAPI counterstaining in non-lesional and lesional skin of patients with psoriasis with (Itch+) and without (Itch–) itch. Immunofluorescence of epidermal KOR expression in frozen skin sections from patients with psoriasis. (A, B) Non-lesional and (C, D) lesional skin with itch (Itch+) (×20 and ×40 magnification, respectively) in comparison with (E, F) non-lesional and (G, H) lesional skin without itch (Itch–) (×20 and ×40 magnification, respectively) documenting decreased KOR expression in lesional psoriatic skin with itch. Western blots using the same KOR antibody (bottom left corner) from non-lesional skin (A) with and (C) without itch in comparison with lesional skin (E) with and (G) without itch. DAPI: (4’,6-diamidino-2-phenylindole).
KOR expression in lesional skin with itch (Fig. 5C, D) was significantly decreased in comparison with lesional skin without itch (p < 0.01; Figs 4c and 5G, H) and the control group (p < 0.001; Figs 4c and 5A, B). However, such KOR expression changes were not observed in non-lesional (Fig. 5E, F) or lesional (Fig. 5G, H) skin without itch compared with the control group (Figs 4c and 1E–G). OPRK1 transcript expression did not present any significant differences between groups with and without itch (Fig. 4d). In addition, there was significant negative correlation between OPRK1 level in non-lesional skin and PASI score (r = –0.70; p < 0.001); no other correlations with studied clinical parameters were found (data not shown).
This study demonstrates that unbalanced expression of ORs (upregulation of the OPRK1/KOR system with stable expression of the OPRM1/MOR system) may contribute to the molecular background of psoriatic itch. Significantly decreased OPRK1 gene expression was found in non-lesional and lesional skin of patients with psoriasis compared with a control group. Moreover, significantly decreased expression of KOR was observed in lesional skin of patients with psoriasis with itch compared with lesional skin without itch, non-lesional skin and control skin. The components of EOS regulate numerous physiological functions and participate in pathways affiliated to psychogenic, neurological, behavioural and stress-related conditions (9, 22, 23). The centrally mediated mechanism of itch and the regulatory role of ORs are known from non-primate and primate models, while human brain imaging studies also support such observations (11–13, 17, 24–26). In the mid-1990s higher levels of ß-endorphin in peripheral blood of patients with psoriasis were noted, suggesting a role for EOS and some neuropeptides in chronic skin neuroinflammation (8). Subsequent studies showed that OPRM1/MOR and OPRK1/KOR are constitutively expressed by epidermal keratinocytes (27). Studies using ORs knockout mice demonstrate that KOR- and MOR-deficient mice present increased epidermal hypertrophy and dysregulation of skin homeostasis (29). The current study supports the idea that peripheral EOS also participates in itch modulation in psoriasis. Tominaga et al. (29) observed that psoralen + ultraviolet A therapy (PUVA) decreased the intensity of itch. This group, for the first time, suggested the downregulation of MOR and restoration of the KOR system in epidermal keratinocytes from PUVA treated patients with atopic dermatitis (28). The same research group reported that patients with psoriasis with and without itch demonstrated unchanged expression of MOR and its endogenous ligand, ß-endorphin, whilst having significantly decreased expression of KOR and dynorphin A in itchy psoriatic skin lesions (21). Molecular biology approaches and animal models have shown that OR-dependent itch pathways are regulated by alternative transcripts and protein variants. While the MOR1 isoform seems to be involved in morphine-induce analgesia, MOR1D is essential for morphine-induced scratching with calcitonin gene-related peptide receptor in heterodimer complexes (30).
The current study found that the OPRK1/KOR system correlates negatively with VAS scoring of itch, which may additionally support the important role of an OPRK1/KOR inhibitory mechanism in itch transmission. Thus, it is possible that topical KOR agonists or MOR antagonists may be helpful in itch therapy (15, 16, 24, 31). Biglardi et al. (32) used topical 1% naltrexone cream on the skin of 13 patients with pruritus, observing an anti-pruritic effect. This observation supports the idea that highly specific opioid ligands, which cannot cross the blood-brain barrier, may be an attractive anti-pruritic therapeutic strategy. Furthermore, nalfurafine hydrochloride, a selective KOR agonist, has demonstrated high effectiveness in different itch subtypes in experimental and clinical trials (16, 33). However, further research is needed into whether unbalanced OR in the CNS–peripheral nervous system axis might result from abnormalities in axonal transport, chronic skin neuroinflammation, social stress-related behaviours or mood disorders, resulting in more frequent somatosensory experiences, such as pain and itch, in psoriasis (7). In contrast to central therapies, in which a high level of adverse side-effects may occur, peripheral opioid pharmacotherapy using highly stereo-specific opioid ligands might be a minimally invasive, but effective, treatment option.
This study was supported financially by a “Preludium” grant from the Polish National Science Center (UMO-2011/01/N/NZ4/04946): “Gene and protein analysis of OPRM1/MOR and OPRK1/KOR opioid receptors expression and neurobiological marker of nerve terminals PGP 9.5 in skin of patients with psoriasis with and without pruritus.” in 2011 to 2013.
D. Nevozhay acknowledges support from programme #1326, of the Ministry of Education and Science, Russian Federation.
The authors declare no conflicts of interest.


 Click to show fullsize
Click to show fullsize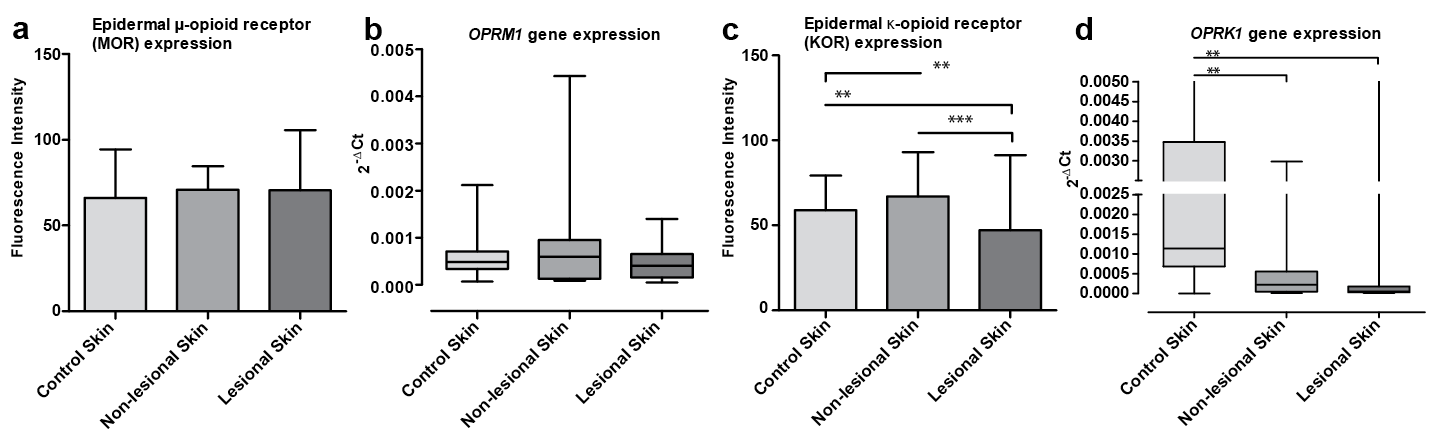 Click to show fullsize
Click to show fullsize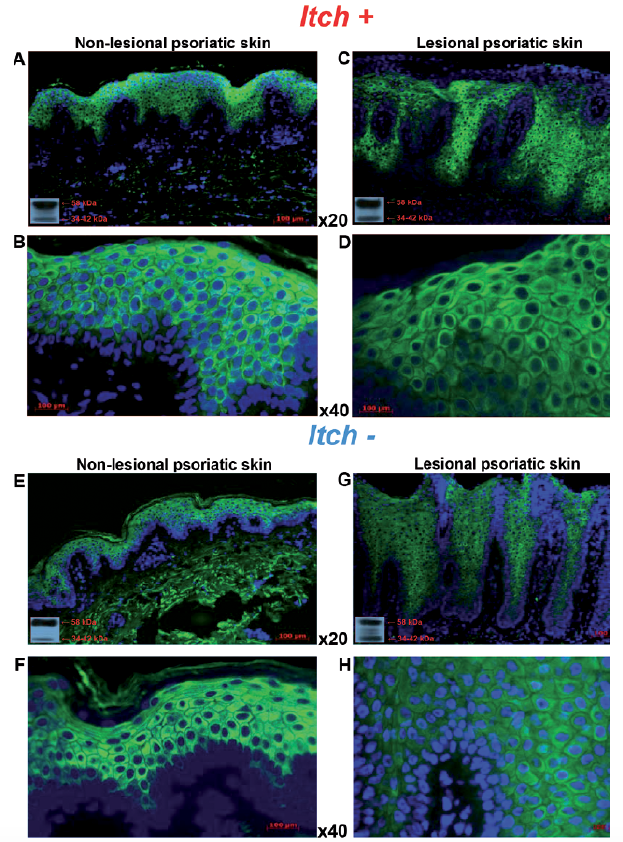 Click to show fullsize
Click to show fullsize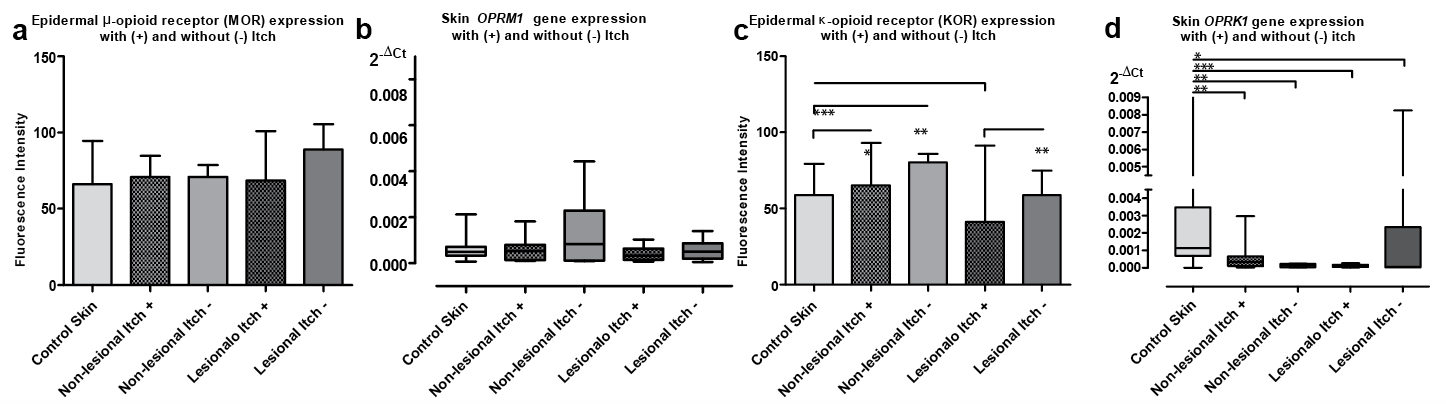 Click to show fullsize
Click to show fullsize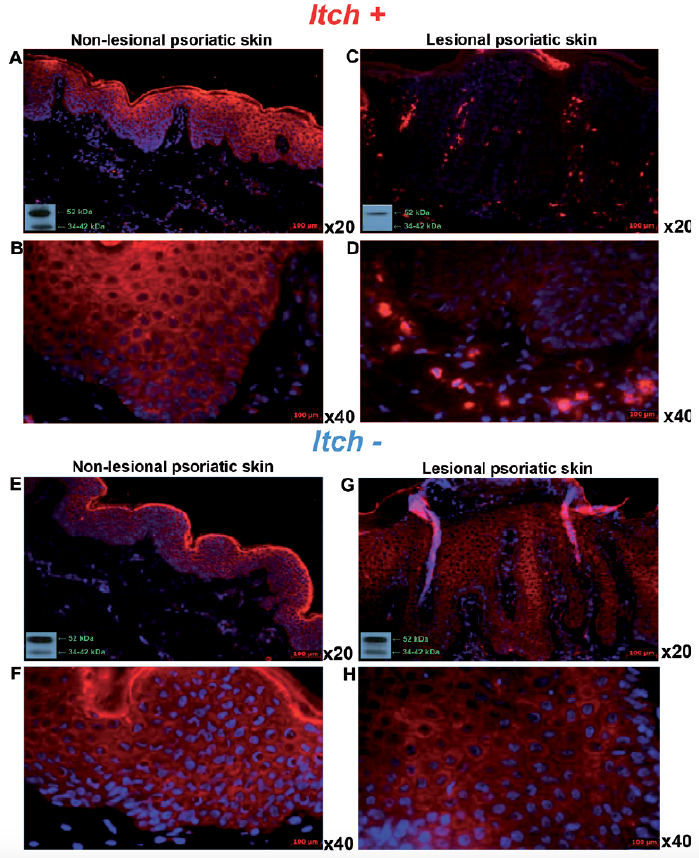 Click to show fullsize
Click to show fullsize