


1Department of Dermatology and Allergy, Herlev and Gentofte Hospital, University of Copenhagen, Hellerup, 2Department of Ophthalmology, 3Eye Pathology Section, Department of Pathology, Rigshospitalet, University of Copenhagen, Copenhagen, Denmark, and 4Coronel Institute of Occupational Health, Academic Medical Center, University of Amsterdam, The Netherlands
Epidermal filaggrin level is affected by ultraviolet B irradiation in animal and experimental models. This study examined the effect of ultraviolet B irradiation on epidermal filaggrin and natural moisturizing factors in vivo in healthy adults (n = 22). Participants were irradiated with 2 minimal erythema doses of ultraviolet B on the skin. Biopsies and tape strips were collected from skin irradiated 24 and 72 h earlier and from non-irradiated skin (control). Real-time quantitative PCR on skin biopsies showed significantly reduced profilaggrin mRNA expression 24 h after irradiation (mean relative mRNA expression ± standard deviation: control, 3.86 ± 2.06 vs. 24 h, 1.52 ± 0.640; p = 0.02; n = 8). Immunohistochemistry showed aberrant spatial distribution of filaggrin protein 72 h after irradiation (n = 3). High-pressure liquid chromatography of tape extracts showed no change in mean total natural moisturizing factor levels after irradiation, but mean trans-urocanic acid was significantly reduced, as expected (n = 8). In conclusion, erythemal doses of ultraviolet B exert acute effects on profilaggrin mRNA and filaggrin protein in human skin in vivo.
Key words: filaggrin; ultraviolet B; acute inflammation; human.
Accepted Mar 27, 2017; Epub ahead of print Mar 30, 2017
Acta Derm Venereol 2017; 97: xx–xx.
Corr: Stine Simonsen, Department of Dermatology and Allergy, Herlev and Gentofte Hospital, Kildegårdsvej 28, DK-2900 Hellerup, Denmark. E-mail: stine.simonsen.01@regionh.dk
Ultraviolet B (UVB) irradiation of human skin (wave-length 280?315 nm) induces numerous biological reactions. The effects are both local and systemic, and vary depending on the dose and type of exposure (1–4). Notably, high-dose UVB irradiation can compromise the functioning of the skin barrier (5, 6).
Filaggrin is an important skin barrier protein. It is formed from processing of profilaggrin encoded by the filaggrin gene (FLG). Monomeric filaggrin emerges in the transition layer between stratum granulosum and stratum corneum (SC). In the SC, it is rapidly degraded to “natural moisturizing factors” (NMFs) including trans-urocanic acid (trans-UCA), a major UVB photon-absorbing chromophore. Trans-UCA is converted to its cis-isomer upon absorption of UVB radiation. Cis-UCA may initiate immunosuppression (7–9).
Approximately 10% of white Europeans are carriers of loss-of-function mutations in FLG; this is a strong genetic risk factor for development of atopic dermatitis (AD).
UVB phototherapy is commonly used to treat inflammatory skin diseases, including AD. It has been shown to restore filaggrin levels in patients with AD after 12 weeks of treatment (10), but little is known about the acute effects of erythemal UVB irradiation on filaggrin.
It is possible that erythemal doses of UVB irradiation suppress expression of profilaggrin mRNA expression and filaggrin protein. This has been investigated in animal models and in human in vitro models. To our knowledge, the present study is the first in vivo study in humans to investigate the acute effect of erythemal UVB irradiation on filaggrin homeostasis.
Twenty-two healthy volunteers of Northern European descent were recruited. Approval of the study was obtained from the local ethics committee (entry numbers H-4-2014-037 and H-4-2011-136). Informed consent was obtained from all participants. All participants were 18 years old or over, and none of them had a history of skin disease or an atopic disposition. Blood samples were collected to determine the presence of the most common FLG mutations in Europe (R501X, 2282del4, and R2447X), and carriers of any of these mutations were excluded. All blood samples were analysed at the Department of Clinical Biochemistry, Herlev and Gentofte Hospital, Hellerup, Denmark. The age and sex of participants and number of participants in each sample group are shown in Table I.
Table I. Demographics (age and sex) and number of participants in each sample group. For each analysis the control samples and ultraviolet B irradiated samples were from the same volunteer
The sources of UVB radiation were 2 Philips TL 20W/12 RS bulbs. These emitted a spectrum of 280?365 nm with a peak emission at about 310 nm (and the main part of the energy having wavelengths between 280 and 320 nm). The irradiation was performed at a distance of 35 cm. Minimal erythema dose (MED) was determined for each participant. Subsequently, 2 MEDs of UVB radiation were administered twice at separate locations on the lower back. The areas of the irradiated spots were 1.0 and 6.0 cm2 (for biopsies and tape stripping, respectively).
Three punch biopsies (4-mm diameter) were collected from 14 volunteers, using local anaesthesia. One biopsy was taken from irradiated skin 24 h after exposure, another from irradiated skin 72 h after exposure, and the last from non-irradiated skin (control). Biopsies were either submerged immediately in liquid nitrogen for mRNA extraction or fixed in 10% formaldehyde for haematoxylin and eosin (H&E) staining and determination of filaggrin expression using immunohistochemistry. Some biopsies were used for both analyses (n = 3). In these cases, the biopsies were immediately divided in 2. One half was submerged in liquid nitrogen and the other half was fixed in 10% formaldehyde.
Biopsy specimens from skin irradiated 24 h previously, 72 h previously, and from non-irradiated skin were used for mRNA extraction. Nucleic acids including total cellular RNA were extracted and purified from tissue samples using the GeneJET RNA Purification Kit according to the protocol supplied by the manufacturer (Thermo Fisher Scientific Biosciences, St Leon-Rot, Germany), at the Department of Clinical Biochemistry, Herlev and Gentofte Hospital, Denmark.
cDNA synthesis and real-time quantitative PCR (RT qPCR) analysis were performed by AROS Applied Biotechnology A/S, Aarhus, Denmark. Briefly, first-strand cDNA synthesis was undertaken using the High Capacity Reverse Transcriptase Kit (Thermo Fisher Scientific Biosciences) using 100 ng total RNA. A specific target amplification was performed using a pool of the 2 PCR assays FLG: Hs00856927_g1 and HPRT1: Hs01003267_m1 (ThermoFisher Scientific), which were also used in the subsequent quantitative PCR. The pre-amplified cDNA and the assays were loaded on a 96.96 GE Chip and run under standard conditions on the Fluidigm Biomark HD system according to the manufacturer’s protocol. Data were analysed using Fluidigm BioMark software ver. 4.1.3 with linear (derivative) baseline correction and the user (detectors) method for Ct threshold settings. Mean Ct values were calculated from the raw data, along with assay standard curve linearity and amplification efficiency.
Biopsy specimens from skin irradiated 24 h previously, skin irradiated 72 h previously, and non-irradiated skin were included in the study.
Immunohistochemical staining was performed as described previously in detail (11). Briefly, sections were incubated with a primary antibody to filaggrin (clone number 15C10, code number NCL-Filaggrin; Novocastra Laboratories Ltd, Newcastle upon Tyne, UK) followed by incubation with a secondary antibody (kit code K5003; Dako A/S, Glostrup, Denmark) Positive controls (human skin tissue) and negative controls (without primary antibody) were included to ensure good antigen retrieval and to prevent false positives. The staining pattern was also characterized. Also, the intensity of staining was scored as negative, weakly positive, moderately positive, or strongly positive. Each specimen was scored blind by 2 investigators (SS and LS). Biopsies were also stained with H&E.
Tape strips of the SC were taken simultaneously from non-irradiated skin and from skin irradiated 24 and 72 h previously. Tape stripping was performed and NMF levels were determined according to the method described previously in detail (12). Briefly, each tape strip was extracted with 0.5 ml 25% (wt/wt) ammonia. The ammonia extract was evaporated and the residue dissolved in 500 μl water before analysis by high-pressure liquid chromatography. The NMF concentrations were normalized according to protein concentration calculated from optical density values determined by SquameScan (CuDerm, Dallas, TX, USA).
L-histidine (HIS), pyrrolidone carboxylic acid (PCA), trans-UCA, and cis-UCA were determined and total NMFs were calculated as the sum.
Statistical analysis was performed with the Wilcoxon matched-pairs signed-rank test (GraphPad Prism version 6.07; GraphPad Software, La Jolla, CA, USA).
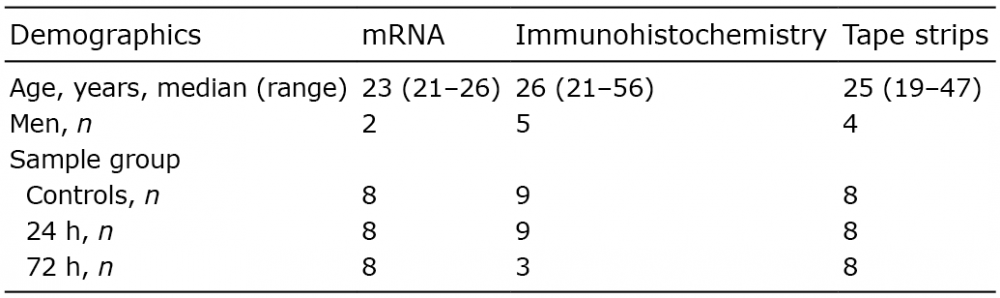
RT qPCR was performed on RNA to determine the level of profilaggrin mRNA in the epidermis in UVB-irradiated and non-irradiated skin (Fig. 1). RT qPCR analysis showed a significant decrease in profilaggrin mRNA 24 h after irradiation compared with control skin (mean relative mRNA expression ± standard deviation (SD): control, 3.86 ± 2.06 vs. 24 h, 1.52 ± 0.640; p = 0.02; n = 8). At 72 h after irradiation, mRNA expression was comparable to the levels found in control skin.
Fig. 1. Profilaggrin mRNA expression in skin biopsies taken from non-irradiated skin (control) and 24 and 72 h after administration of a single dose of 2 minimal erythema doses (n = 8). Data are expressed as relative expression of FLG compared with the house keeping gene HPRT1. The columns represent mean and the error bars represent standard deviation.
Twenty-four hours after UVB exposure, irradiated skin had several suprabasal sunburn cells, pyknotic nuclei, and eosinophilic cytoplasm (n = 9). Seventy-two hours after UVB exposure, the sunburn cells had moved upwards and were found in conjunction with parakeratosis in a layer beneath normal-appearing SC and above a markedly hyperplastic stratum granulosum (Fig. 2) (n = 3).
Fig. 2. Histopathological changes caused by 2 minimal erythema doses (MEDs) of ultraviolet B (UVB) irradiation demonstrated by haematoxylin-eosin (H&E) staining and filaggrin immunostaining. (A) Non-irradiated skin showing normal epidermis (H&E staining, magnification ×200). (B) Skin 24 h after UVB irradiation showing sunburn cells in suprabasal layers (arrow) (H&E staining, magnification ×200) (C) Skin 72 h after UVB irradiation showing hyperplasia, parakeratosis (arrow) and sunburn cells (H&E staining, magnification ×200). (D) Non-irradiated skin showing normal filaggrin expression (magnification ×200). (E) Skin 24 h after irradiation showing no changes in filaggrin expression (magnification ×200). (F) Skin 72 h after UVB irradiation showing 2 zones of filaggrin positive cells (arrows) (magnification ×200).
Filaggrin protein was examined in irradiated and non-irradiated skin using immunohistochemical staining. A large degree of inter-individual variation was found among the subjects included, as judged by staining intensity and extension of positive staining into the epidermis. No change was found in staining pattern or overall staining intensity 24 h after UVB irradiation (Fig. 2E). In all 3 biopsies taken 72 h after irradiation, 2 filaggrin zones were observed: an upper zone located further up in the SC compared with non-irradiated skin where the filaggrin-positive cells appeared to be less nucleated, and a lower zone of filaggrin-positive cells located below the layer showing sunburn cells and parakeratosis. In this lower zone, the filaggrin-positive cells resembled stratum granulosum cells. In addition, the upper filaggrin zone appeared thinner (Fig. 2F).
To investigate whether UVB irradiation could affect filaggrin degradation products, we evaluated tape strips from non-irradiated and irradiated skin. As expected, the mean trans-UCA level was significantly reduced 24 h after irradiation (control, 0.095 ± 0.029 mmol/g tissue vs. 24 h, 0.043 ± 0.014 mmol/g; p < 0.01; n = 8) (Fig. 3A).
Fig. 3. (A) Mean trans-urocanic acid (UCA), (B) mean cis-UCA, (C) mean total natural moisturizing factor (NMF) (i.e. L-histidine, pyrrolidone carboxylic acid, trans-UCA, and cis-UCA), (D) mean pyrrolidone carboxylic acid (PCA), and (E) mean L-histidine (HIS), in non-irradiated skin (Control) and 24 h and 72 h after administration of a single dose of two minimal erythema doses (MEDs) (n = 8). Error bars represent standard deviation (SD). *Significant difference compared with controls.
A similar decrease was seen at 72 h after irradiation. Correspondingly, cis-UCA was increased at 24 and 72 h after irradiation (Fig. 3B). There was no difference between non-irradiated and irradiated skin in mean total NMF (Fig. 3C), mean PCA (Fig. 3D) or mean HIS (Fig. 3E) levels.
This study showed that profilaggrin mRNA expression is transiently reduced when human skin is exposed to 2 MEDs of UVB irradiation. Furthermore, we found distinct morphological changes and aberrant spatial distribution of filaggrin protein in the epidermis 72 h after exposure.
In contrast to our results, a recent in vitro study found that profilaggrin mRNA expression was increased 24 h after narrowband UVB irradiation (13). The inconsistency with our data may be explained by differences in study design. Firstly, the UVB dose used in the in vitro study was performed with a much lower dose of UVB that did not cause sunburn cells. Secondly, that study used a human epidermal model based only on keratinocytes. Thus, dermis and vasculature were absent and there were no immune cells.
It is likely that profilaggrin mRNA expression is reduced because of DNA damage and inflammation induced by the UVB irradiation. Thus, tumour necrosis factor-α, which is upregulated as early as 6 h after UVB irradiation of human skin (14), inhibits profilaggrin mRNA expression in primary human keratinocytes in vitro (15) and may also contribute to the downregulation of profilaggrin mRNA. No other UVB-induced cytokines have been investigated in this context, but numerous AD-associated cytokines have been shown to downregulate FLG mRNA expression and filaggrin protein expression in vitro (16, 17). We have not validated the stability of the reference gene HPRT1 in our experimental setup. HPRT1 has shown some instability following UVB irradiation of cultured keratinocytes (18). Our data should therefore be interpreted with caution.
Our immunohistochemical staining showed a markedly altered spatial distribution of filaggrin 72 h after irradiation with 2 MEDs of UVB. In vitro and xenograft studies have failed to find similar changes (19, 20), but our data are in agreement with the results of an in vivo study of guinea pig skin (21). The results from the latter study also showed 2 filaggrin-positive zones 72 h after irradiation with 2 MEDs. The authors mainly focused on the impact of chemically induced inflammation on filaggrin expression, and the morphology of the UVB-irradiated skin was not described in detail. However, they concluded that UVB induces the keratinocytes to skip the granular phase at 24 h after exposure and consequently produce a layer of filaggrin-free SC, which we do not confirm. Moreover, at 72 h after irradiation we found that the lower filaggrin zone was aberrantly present in stratum granulosum.
To our knowledge, no other studies have investigated the immediate change in NMF levels after erythemal UVB irradiation. NMF levels are dependent on the amount of filaggrin protein in the epidermis, the rate of filaggrin breakdown, and possibly also the rate of desquamation. The isomerization of trans-UCA to cis-UCA in the present study did not influence the total NMF level. The reason that we found no changes in total NMF levels at 24 and 72 h after exposure despite decreased filaggrin expression was probably caused by the fact that the SC samples originated from the upper layers of the SC. The layers with reduced levels of filaggrin due to UV exposure would need at least 2 weeks to reach more superficial SC layers. In vitro human studies and in vivo animal studies do not necessarily represent human responses in vivo accurately, and our data provide more insight into the acute effects of UVB exposure. We used MED test to ensure that all participants were exposed to equal biological doses of UVB irradiation. This was done to achieve maximum comparability between participants because administration of the same level of UVB irradiation in joules/cm2 could cause inter-individual differences in erythema, inflammation, and tissue damage due to individual variation in UVB sensitivity. The presence of sunburn cells, the significant decrease in trans-UCA, and the presence of erythema of the irradiated skin confirm that the UVB irradiation was administered correctly. However, only healthy white Northern Europeans were included, and our data may not apply to individuals with skin conditions or to other ethnic groups. Elucidation of the immediate effect of UVB irradiation on filaggrin in the skin and therefore the possible effect on the skin barrier is important for our understanding of the risk of eczema and flare of eczema. Notably, patients with AD with acute skin inflammation often cannot tolerate UVB irradiation at the start of treatment and many patients with AD complain of worsening of dermatitis following start-up of UVB treatment. Furthermore, reduction in filaggrin levels and thereby trans-UCA or trans-UCA levels directly may lead to increased penetration of UVB irradiation (22), which could contribute to a higher risk of cancer. Patients with AD often have lower filaggrin levels in the skin and possibly an increased risk of basal cell carcinoma (23, 24). Furthermore, a small study has shown an increased risk of squamous cell carcinoma in individuals with homozygous FLG loss-of-function mutations (25), but further studies are needed.
In conclusion, we found that UVB irradiation has a significant impact on profilaggrin mRNA synthesis in human skin in vivo and we have confirmed that, after irradiation, there is a distinct alteration in the location of filaggrin protein.
The authors would like to thank Anne Falensteen Lauritzen (Senior Consultant) of the Department of Pathology, Gentofte and Herlev Hospital, for expert help with interpretation of histology.
This work was supported by grants from the Capital Region of Denmark Foundation for Health Research, the Aage Bang Foundation, and the Jacob and Olga Madsen Foundation. Stine Simonsen was supported by a grant from Copenhagen University.


 Click to show fullsize
Click to show fullsize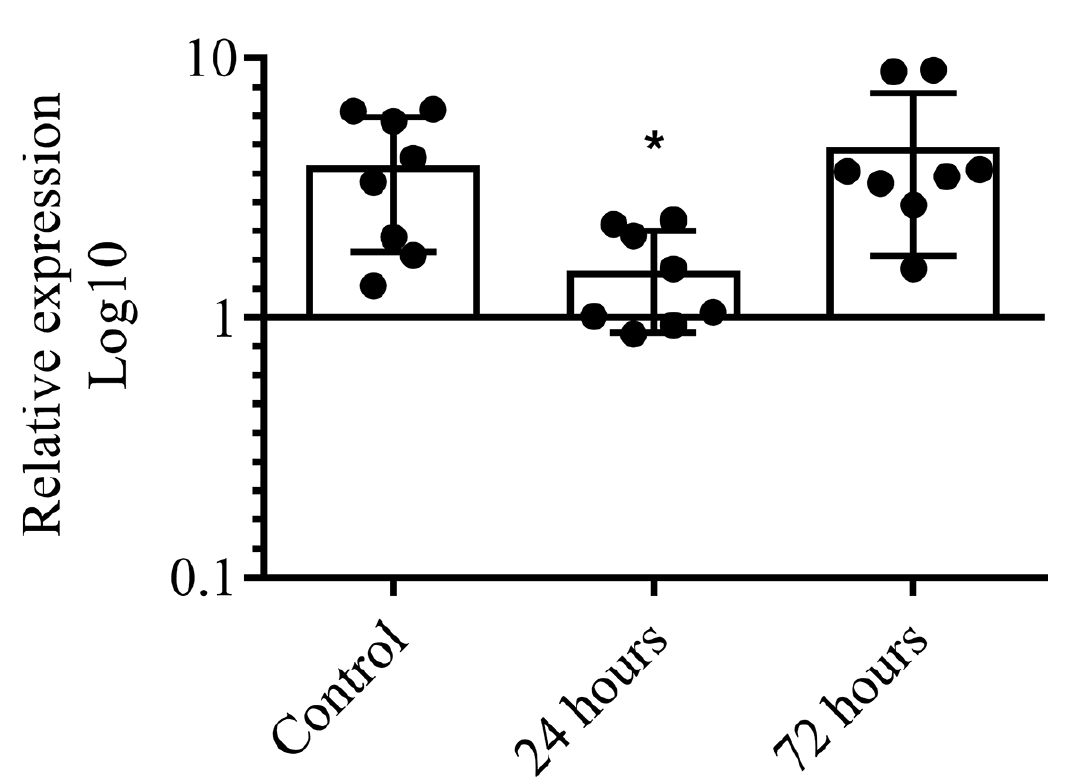 Click to show fullsize
Click to show fullsize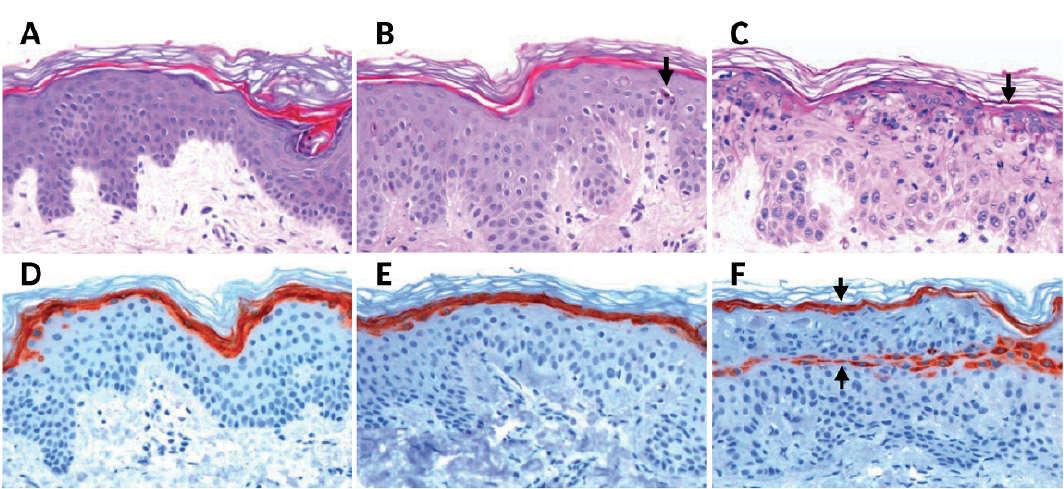 Click to show fullsize
Click to show fullsize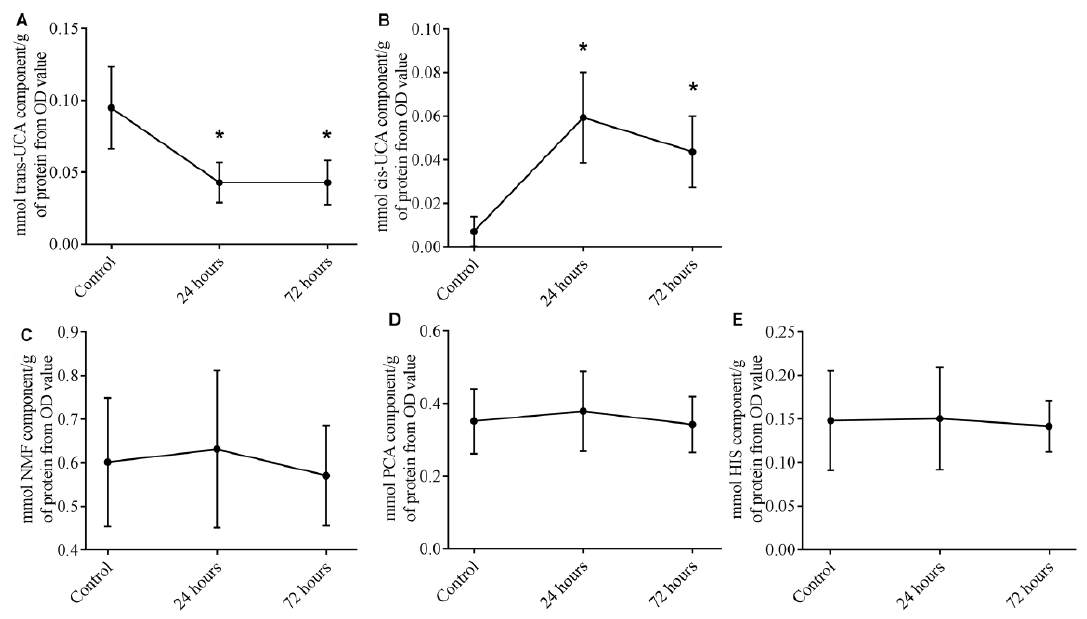 Click to show fullsize
Click to show fullsize