


Department of Dermatology, University of Tokyo Graduate School of Medicine, Tokyo, Japan
Interleukin-19 (IL-19), a pro-inflammatory cytokine known to stimulate the production of T helper type 2 (Th2) cytokines, is induced by IL-17A and highly expressed in the lesional skin of psoriasis and atopic dermatitis (AD). This aim of this study was to investigate whether IL-19 is involved in cutaneous T-cell lym-phoma (CTCL) and AD. IL-19 levels were significantly higher in the sera of patients with AD and those with advanced-stage CTCL than in normal controls, correlating significantly with clinical disease markers. IL-19 mRNA levels in lesional skin of both diseases were significantly elevated. Immunohistochemical staining revealed that IL-19 was expressed in the epidermis of AD skin and CTCL skin. In vitro, IL-17A and IL-4 increased IL-19 mRNA expression in human keratinocytes. Thus, IL-19 was increased in the sera and skin of AD and CTCL. These results suggest that IL-19 is important for bridging Th17 to Th2 in these diseases.
Key words: IL-19; atopic dermatitis; cutaneous T-cell lymphoma; keratinocytes; IL-4; IL-17A.
Accepted Jun 8, 2017; Epub ahead of print Jun 9, 2017
Acta Derm Venereol 2017; 97: xx–xx.
Corr: Makoto Sugaya, Department of Dermatology, the University of Tokyo Graduate School of Medicine, 7-3-1 Hongo, Bunkyo-ku, Tokyo 113-8655, Japan. E-mail: sugayam-der@h.u-tokyo.ac.jp
Interleukin (IL)-19 is a member of the so-called IL-10 family of cytokines. IL-19 was discovered in 2000 by means of a bioinformatics search for IL-10-related molecules (1). Important cellular sources of IL-19 are thought to be monocytes (1–3), keratinocytes (4–8) and bronchial epithelial cells (9). IL-4 and IL-17A have been reported to induce IL-19 expression by human keratinocytes (4, 5, 7, 8). The IL-19 receptor consists of IL-20R1 and IL-20R2 (10–12). The IL-19 subfamily has 3 members, IL-19, IL-20 and IL-24, which have a very similar tertiary structure (13) and are able to bind to and signal through IL-20R1 and IL-20R2. Keratinocytes express these receptors, suggesting that IL-19 can act in an autocrine manner (8).
Many groups have documented increased IL-19 expression in the diseased skin of psoriasis in comparison with non-diseased skin of the same patients or that of healthy controls (4, 5, 14–16). IL-19 is also reported to be up-regulated in lesional skin of atopic dermatitis (AD) (4, 5). AD is a chronic inflammatory skin disease that affects 10–20% of children and 1–3% of adults in developed countries. It is well known that T helper type 2 (Th2) cells, expressing IL-4, IL-5, and IL-13, play an essential role in the pathophysiology of AD (17). IL-19 has been demonstrated to increase Th2 cytokine expression in activated T cells (3, 18). Serum IL-19 levels are elevated in patients with asthma, suggesting an essential role in the pathogenesis of asthma (18), which is another Th2 cell-mediated disease. Serum IL-19 levels in patients with AD, however, have not been investigated previously.
Cutaneous T-cell lymphoma (CTCL) is a malignancy of skin-trafficking T cells. Mycosis fungoides (MF) and Sézary syndrome (SS) are the most common types of CTCL. Prior studies have shown increased levels of Th2 cytokines and Th2-associated genes in T cells from patients with CTCL (19). Although up-regulated IL-19 in breast cancer has been shown to promote tumour progression (20), little is known about an association between IL-19 and other malignancies. IL-19 has not been investigated for possible involvement in CTCL. The aim of this study was therefore to examine serum IL-19 levels and IL-19 expression in lesional skin of AD and CTCL. A further aim was to investigate which cytokines would induce IL-19 expression by normal human keratinocytes.
Messenger RNA (mRNA) was obtained from biopsy materials of lesional skin of MF/SS (n = 17: patch MF 4, plaque MF 3, tumour MF 5, erythrodermic MF 2, SS 3; mean ± standard deviation (SD) age: 57.6 ± 12.7 years; 12 males and 5 females), AD (n = 7; mean ± SD age: 36.3 ± 16.0 years; 5 males and 2 females), and normal skin adjacent to benign skin tumours (n = 8; mean ± SD age: 49.1 ± 21.0 years; 4 males and 4 females) using RNeasy Fibrous Tissue Mini Kit (QIAGEN, Valencia, CA, USA). Samples for immunohistochemistry were lesional skin of MF/SS (n = 10: patch MF 2, plaque MF 3, tumour MF 3, SS 2) or AD (n = 10), and normal skin adjacent to benign skin tumours (n = 10). Serum samples were obtained from 28 patients with MF/SS (patch MF 7, plaque MF 10, tumour MF 9, erythrodermic MF 1, SS 1; mean ± SD age: 54.5 ± 18.1 years; 15 males and 13 females), 35 patients with AD (10 mild cases, 10 moderate cases, and 15 severe cases; mean ± SD age: 30.8 ± 12.0 years; 23 males and 12 females), and 17 healthy control subjects (mean ± SD age: 47.6 ± 20.6 years; 11 males and 6 females). The healthy controls had no history of allergy, AD, psoriasis, or CTCL. All samples were collected during daily clinical practice in the University of Tokyo Hospital. The medical ethics committee of the University of Tokyo approved all described studies and the study was conducted according to the principles of the Declaration of Helsinki. Informed consent was obtained to use blood and skin samples from patients and healthy controls. All patients with MF/SS were given diagnoses according to WHO classification for cutaneous lymphomas (21). All patients with AD were enrolled according to the criteria of Hanifin & Rajka (22). In patients with CTCL, clinical stages IA, IB and IIA were defined as early stage and stages IIB, IIIA, IIIB, IVA and IVB were defined as advanced stage.
Normal human epidermal keratinocytes (NHEK) were purchased from Kurabo Industries (Osaka, Japan). NHEK were cultured in 75 cm2 cell culture flasks (Sigma-Aldrich, St Louis, MO, USA) at 37°C, 5% CO2 in Humedia-KB2 (Kurabo Industries) supplemented with Human Keratinocyte Growth Supplement sets (Kurabo Industries). The cells received fresh medium every 3 days and were subcultured every 10 days. When 80% confluence was achieved, the cells were trypsinized, washed, and resuspended in the medium at 5×105 cells/ml, and 1 ml was added to each well of the 12-well plates (Becton Dickinson Labware, Franklin Lakes, NJ, USA). When the cells reached confluence, the medium was completely removed and 1 ml serum-free medium was added to each well. Simultaneously, recombinant human IL-4 (R&D Systems, Minneapolis, MN, USA) IL-13 (BioLegend, San Diego, CA, USA), IL-17A (R&D Systems), IL-22 (R&D Systems) and IFN-γ (R&D Systems) were added, and the cells were incubated at 37°C and 5% CO2. The concentration of IL-17A was 1, 10, or 100 ng/ml, IL-4 and IL-22 were 10 or 100 ng/ml, IL-13 was 0.2, 2, or 20 ng/ml, and IFN-γ was 0.1, 1 or 10 ng/ml. After 24 h, cells were processed by TRIzol Reagent (Invitrogen, Carlsbad, CA, USA) for isolation of total mRNA, according to the manufacturer’s instructions.
Total mRNA was obtained from human skin samples with RNeasy Fibrous Tissue Mini Kit (QIAGEN) and from cells with TRIzol Reagent (Invitrogen). Complementary DNA was synthesized using ReverTra Ace® qPCR RT Master Mix (TOYOBO, Osaka, Japan). mRNA levels were analysed using real-time quantitative PCR (RT-PCR) with THUNDERBIRD® SYBR® qPCR Mix (TOYOBO) on an ABI Prism 7000 sequence detector (Applied Biosystems, Foster City, CA, USA). Primers for human GAPDH, IL-19, IL-17A, and IL-4 were as follows: human GAPDH forward, 5’-ACC CAC TCC TCC ACC TTT GA -3’ and reverse, 5’-CAT ACC AGG AAA TGA GCT TGA CAA-3’; human IL-19 forward, 5’-AAG GAT CAT CAG GAG CCA AA-3’ and reverse, 5’-CAC TGC CTC TGT TCC TGA CA-3’; human IL-17A forward, 5’-ACT ACA ACC GAT CCA CCT CAC-3’ and reverse, 5’-ACT TTG CCT CCC AGA TCA CAG-3’; human IL-4 forward, 5’-CAC AGG CAC AAG CAG CTG AT-3’ and reverse, 5’-CTC TGG TTG GCT TCC TTC ACA-3’. All samples were analysed in parallel for GAPDH gene expression as an internal control. The relative expression levels of each gene were determined by the 2–ΔΔCT method.
Immunohistochemical staining for IL-19 was performed in normal skin adjacent to benign skin tumours, as controls (n = 10), and lesional skin of patients with AD (n = 10), CTCL (n = 10), and psoriasis (n = 10). Briefly, 5-μm thick tissue sections from formaldehyde-fixed and paraffin-embedded samples were de-waxed and rehydrated. These sections were then stained with rabbit anti-human IL-19 polyclonal Ab (Abcam, Cambridge, UK), followed by ABC staining (Vector Lab, Burlingame, CA, USA). Diaminobenzidine was used for visualizing the staining, and counterstaining with Mayer haematoxylin was performed, according to the manufacturer’s instructions. The positivity of staining was ranked from negative to strong positive.
Immunoreactive IL-19 in sera was quantified by human enzyme-linked immunosorbent assay (ELISA) kits (R&D Systems). These assays employ the quantitative sandwich enzyme immunoassay technique. Optical densities were measured at 450 nm, with the correction wavelength set at 570 nm, using a Bio-Rad Model 550 microplate reader (Bio-Rad Laboratories, Hercules, CA, USA). The concentrations were calculated from the standard curve generated by a curve-fitting program according to the manufacturer’s instructions.
Statistical analysis was performed using Prism Version 5 software (GraphPad, San Diego, CA, USA). Significant differences were analysed with the Mann–Whitney U test. Correlation coefficients were determined using Spearman’s rank correlation test. p-values < 0.05 were considered statistically significant.
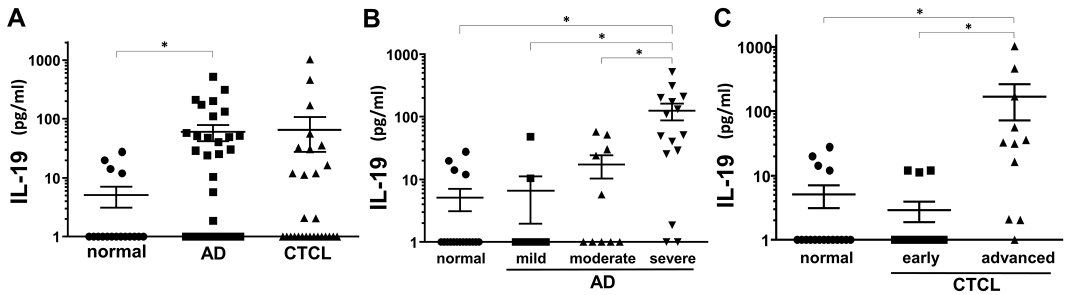
To investigate IL-19 involvement in AD and CTCL, we first examined IL-19 protein levels in sera of patients with AD and CTCL. IL-19 protein levels in sera of AD patients were significantly elevated compared with those of normal controls (Fig. 1A). Serum IL-19 levels increased according to AD severity (Fig. 1B). IL-19 protein levels in sera of patients with CTCL tended to be increased, although there was no statistical significance (Fig. 1A). When divided by clinical stages, patients with advanced stage CTCL showed significant elevation in serum IL-19 levels compared with normal controls and patients with early stage CTCL (Fig. 1C). Thus, serum IL-19 levels were elevated in AD and patients with advanced stage CTCL.
Fig. 1. Serum IL-19 protein levels in patients with atopic dermatitis (AD) or cutaneous T-cell lymphoma (CTCL), and normal controls. The measured values from individual patients were plotted by dots. Error bars represent standard errors of the mean. *p < 0.05. (A) Serum interleukin-19 (IL-19) protein levels in AD (n = 35), CTCL (n = 28), or normal controls (n = 17). (B) Serum IL-19 protein levels in mild (n = 10), moderate (n = 10), or severe (n = 15) AD patients and normal controls (n = 17). (C) Serum interleukin (IL)-19 protein levels in early (n = 17) or advanced (n = 11) stage cutaneous T-cell lymphoma (CTCL) patients and normal controls (n = 17). As for patients with CTCL, clinical stage IIB or more were defined as advanced stage and clinical stage I and IIA were defined as early stage.
The association between serum IL-19 levels and clinical disease markers was examined. Eczema Area and Severity Index (EASI) score, a severity index of AD, was positively associated with serum IL-19 levels in patients with AD (Fig. 2A). More-over, serum levels of thymus and activation-regulated chemokine (TARC) and the number of eosinophils in peripheral blood significantly correlated with serum IL-19 levels in patients with AD (Fig. 2B, C). Serum IL-19 levels were positively associated with serum soluble IL-2 receptor (sIL-2R) levels in patients with CTCL (Fig. 2D). Thus, serum IL-19 levels positively correlated with clinical disease markers of AD and CTCL.
Fig. 2. Correlation between serum interleukin-19 (IL-19) protein levels and clinical markers in atopic dermatitis (AD) and cutaneous T-cell lymphoma (CTCL). (A) Correlation between serum IL-19 protein levels and Eczema Area and Severity Index (EASI) in AD (n = 21). (B) Correlation between serum IL-19 levels and serum thymus and activation-regulated chemokine (TARC) levels in AD (n = 29). (C) Correlation between serum IL-19 levels and the number of eosinophils in AD (n = 33). (D) Correlation between serum IL-19 levels and serum soluble IL-2 receptor (sIL-2R) levels in CTCL (n = 23).
Levels of expression of IL-19, IL-4 and IL-17A mRNA in normal skin and lesional skin of AD and CTCL were examined. IL-19 mRNA levels were significantly elevated in both AD and CTCL skin compared with those of normal skin (Fig. 3A). In AD skin, IL-19 mRNA expression levels positively correlated with IL-4 mRNA levels (Fig. 3B). In CTCL skin, IL-19 mRNA levels positively correlated with both IL-4 and IL-17A mRNA levels (Fig. 3C, D). Thus, IL-19 mRNA expression levels were elevated in AD and CTCL skin, which correlated with IL-4 and IL-17A mRNA expression levels.
Fig. 3. Interleukin-19 (IL-19) mRNA expression in lesional skin of atopic dermatitis (AD) or cutaneous T-cell lymphoma (CTCL), and normal control skin and that of human epidermal keratinocytes stimulated with cytokines. (A) IL-19 mRNA expression in lesional skin of AD (n = 7) or CTCL (n = 17) and normal skin (n = 8). The measured values from individual patients were plotted by dots. Error bars represent standard errors of the mean (SEM). *p < 0.05. (B) Correlation between IL-19 and IL-4 mRNA levels in AD lesional skin (n = 7). (C) Correlation between IL-19 and IL-4 in CTCL lesional skin (n = 17). (D) Correlation between IL-19 and IL-17A in CTCL lesional skin (n = 17). (E) IL-19 relative mRNA levels in human epidermal keratinocytes stimulated with IL-17A (1, 10, 100 ng/ml) or medium control for 24 h. The mean level in medium control was arbitrarily set to 1 and all values represent the mean+SEM (n = 6). All experiments were done twice and representative results are shown. *Indicates significant difference compared with medium control. *p < 0.05 by Mann–Whitney U test. (F) IL-19 relative mRNA levels in human epidermal keratinocytes stimulated with IL-4 (10, 100 ng/ml) or medium control for 24 h. The mean level in medium control was arbitrarily set to 1 and all values represent the mean+SEM (n = 6). All experiments were done twice and representative results are shown. *Indicates significant difference compared with medium control. *p < 0.05.
To study regulation of IL-19 expression by keratinocytes, IL-19 mRNA levels were investigated with quantitative RT-PCR using NHEK stimulated with IL-17A, IL-4, IL-13, IL-22, IFN-γ, or medium only. IL-17A increased IL-19 mRNA expression by NHEK in a dose-dependent manner (Fig. 3E). Stimulation with IL-4 also significantly increased IL-19 mRNA levels, although the levels of increase were much smaller than IL-17A stimulation (Fig. 3F). Stimulation with IL-13, IL-22, or IFN-γ did not change IL-19 mRNA expression (data not shown). Thus, IL-19 mRNA expression by human keratinocytes increased by IL-17A or IL-4 stimulation, which was consistent with previous reports (4, 5, 7, 8).
Immunohistochemical staining of IL-19 revealed that epidermis in AD skin strongly expressed IL-19 (Fig. 4). Keratinocytes in CTCL lesional skin were slightly positive for IL-19 (Fig. 4). Lesional skin of psoriasis was also stained positive, as reported previously (16, 23). The immunohistochemistry of 10 cases in each group are summarized in Table I. Thus, epidermis of AD, CTCL and psoriasis lesional skin expressed IL-19.
Fig. 4. Immunohistochemistry of interleukin-19 (IL-19) in normal skin and lesional skin of atopic dermatitis (AD), cutaneous T-cell lymphoma (CTCL) or psoriasis.
Table I. Interleukin-19 expression in lesional skin of normal skin, atopic dermatitis (AD), cutaneous T-cell lymphoma (CTCL) and psoriasis
This study showed high levels of IL-19 expression in the sera and lesional skin of patients with AD and CTCL. In patients with AD and CTCL, serum IL-19 levels positively correlated with clinical disease markers. In lesional skin, IL-19 mRNA levels positively correlated with IL-4 mRNA levels in AD, and with IL-4 and IL-17A mRNA levels in CTCL. In vitro, IL-17A and IL-4 increased IL-19 expression by human keratinocytes. Immunohistochemical staining revealed that epidermis in AD, CTCL, and psoriasis lesion skin expressed IL-19. Taken together, IL-19 is highly expressed in CTCL as well as AD, suggesting a possible role in the development of these diseases.
IL-19 expression was increased at both mRNA and protein levels in AD lesional skin (see Figs 3A and 4), which was consistent with previous reports (4). CTCL lesional skin, whose cytokine/chemokine profiles are similar to AD skin (24), also expressed IL-19 (Figs. 3A and 4). Moreover, we first showed serum IL-19 levels were significantly increased in both diseases, positively correlating with the clinical severity (Fig. 1) and other disease markers (Fig. 2). EASI, serum TARC levels, and the number of eosinophils clearly indicate AD severity (25–27) and serum sIL-2R levels reflect CTCL disease activity (28). The role of IL-19 has been proposed in psoriasis, but our results suggest that IL-19 is also involved in AD and CTCL.
IL-19 mRNA levels positively correlated with IL-4 mRNA levels in AD and CTCL skin (Figs. 3B and C). Previous studies reported that IL-19 induced Th2 cytokines (3, 18) and that IL-19 expression was up-regulated by Th2 cytokines (5, 29). Serum IL-19 levels were up-regulated in patients with asthma, a representative Th2 disease (18). Our results are consistent with the fact that AD and CTCL at advanced stage are both Th2-dominant diseases.
Both IL-4 and IL-17A have been reported to up-regulate IL-19 expression by keratinocytes (4, 5, 8). In this study, stimulation with IL-17A significantly enhanced IL-19 mRNA expression by human keratinocytes in a dose-dependent manner (Fig. 3E). Stimulation with IL-4 also increased IL-19 mRNA expression, although the levels of increase were much smaller than with IL-17A stimulation (Fig. 3F). Interestingly, IL-19 mRNA levels positively correlated with both IL-17A mRNA levels and IL-4 mRNA levels in CTCL (Fig. 3C, D), and IL-19 mRNA levels positively correlated with IL-4 mRNA levels in AD (Fig. 3B). IL-19 expression by monocytes was reported to be increased by lipopolysaccharide stimulation, but not by IL-4 (1). Since IL-19 expression by keratinocytes was increased by IL-17A and IL-4, which are abundantly expressed in inflammatory skin conditions and CTCL, and significant positive correlation between IL-19 mRNA levels and those of IL-17A or IL-4 was detected in CTCL and AD skin, keratinocytes seem to be the main source of IL-19 in these diseases. Immunohistochemical staining of IL-19 was also supportive.
Although Th2-dominancy is well known in AD, some studies have indicated that IL-17A was also involved in the pathogenesis of AD (30–33). In addition, IL-17 expression has been reported in lesion skin from patients with CTCL (34, 35). As IL-19, which is strongly induced by IL-17A (Fig. 3E), can increase Th2 cytokine expression in T cells (3, 18), IL-19 may function as a bridge from Th17 to Th2. As IL-19 is also induced by IL-4, IL-19 seems to be commonly expressed in various diseases and to serve as a multi-functional player. Because AD and CTCL have different characteristics, such as relatively low levels of antimicrobial peptide expression and a high susceptibility to infection, compared with psoriasis (36, 37), the function of IL-19 in the former two diseases may be different from that in psoriasis due to different microenvironment. Although further study is necessary to elucidate the underlying mechanism, it is noteworthy that IL-19 is also involved in AD and CTCL.
In summary, IL-19 was up-regulated in the sera and skin of both AD and CTCL. IL-19 expression in keratinocytes was up-regulated by IL-17A and IL-4. IL-19 may be important for bridging Th17 to Th2 in AD and CTCL.
The authors would like to thank Tamami Kaga for technical assistance.
The authors declare no conflicts of interest.


 Click to show fullsize
Click to show fullsize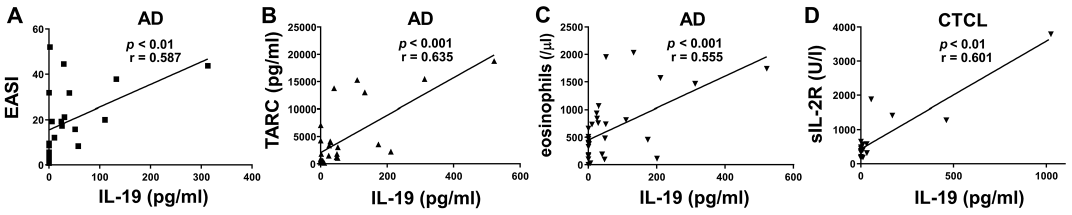 Click to show fullsize
Click to show fullsize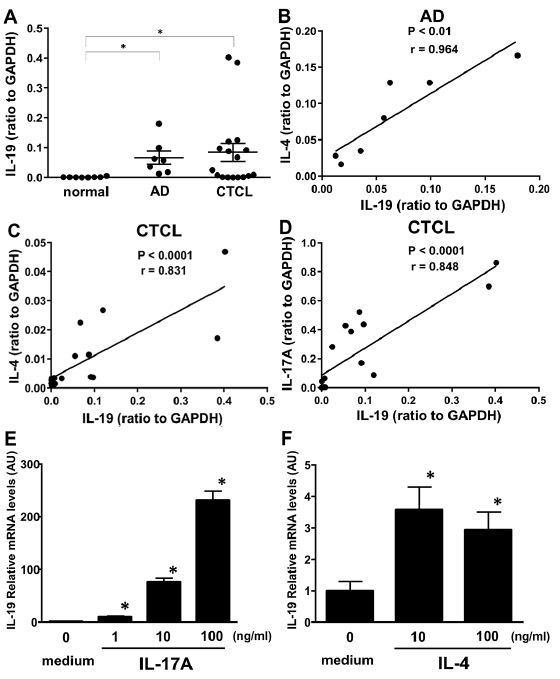 Click to show fullsize
Click to show fullsize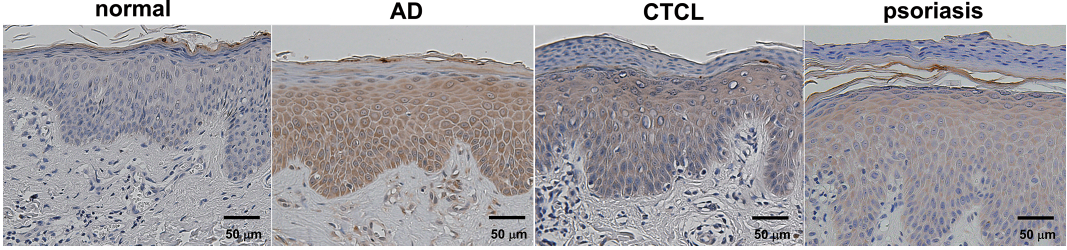 Click to show fullsize
Click to show fullsize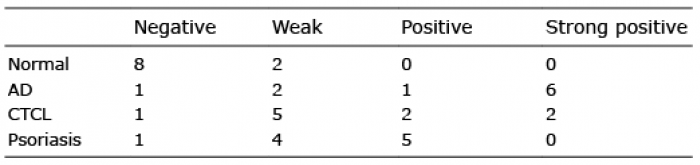 Click to show fullsize
Click to show fullsize