


1Department of Psychosomatics and Psychotherapy, 2Center for Chronic Pruritus (KCP), 3Department of Clinical Radiology and 5Department of Dermatology, University of Münster, Münster, and 4Department of Anesthesiology and Intensive Care Medicine, Karl Feuerstein Professorship, Medical Faculty Mannheim, University of Heidelberg, Mannheim, Germany
The central processing of itch is not completely understood. This is the first study to use functional magnetic resonance imaging (fMRI) to examine the central modulation by distraction of experimentally induced itch. A total of 33 healthy volunteers were examined with fMRI. Periods of itch induction without distraction and itch with distraction by a Stroop task (psychological test, where the participants have to decide if the colour of the writing corresponds to the written word, for example if “red” is written in red or not) were counterbalanced during the scanning to examine task-specific changes in blood oxygenation level dependent-signal. The intensity of the subjects’ itch sensation, desire to scratch and pain sensation were evaluated. Distraction by a Stroop task did not reduce itch intensity or urge to scratch. However, the Stroop task led to significantly higher activation of the left brainstem when it followed the “pure” itch sensation. Itch and pain seem to have similar inhibition pathways, particularly concerning brainstem activation during distraction. But as itch sensation, in contrast to pain, could not be sufficiently reduced by distraction, both entities might have different modulation systems.
Key words: fMRI; itch; scratch; pain; pain inhibition system.
Acta Derm Venereol 2017; 97: xx–xx.
Accepted Jun 22, 2017; Epub ahead of print Jun 27, 2017
Corr: Astrid Stumpf, Department of Psychosomatics and Psychotherapy, University of Münster, Domagkstrasse 22, DE-48149 Münster, Germany. E-mail: astrid.stumpf@ukmuenster.de
Itch and pain are unpleasant sensory experiences that have many characteristics in common. Paralleling clinical pain (1, 2), itch leads to a huge reduction in quality of life and thus can provoke comorbidities such as anxiety disorders or depression (3–5). Like chronic pain, the treatment of chronic itch is often insufficient and represents a challenge for the physician (6–9).
During recent years, our knowledge of central processing and modulation of itch has increased. Positron emission tomography (PET) and functional magnetic resonance imaging (fMRI) during experimental itch stimulation have revealed activation of brain regions responsible for motor planning, sensory aspects and emotional processes (10–20). The activation patterns of itch and pain overlap to a great extent. However, relative increases in different brain areas may differ. For example, itch-induced activation of the somatosensory areas SI and SII appears to be weaker, whereas activation of motor areas is stronger (21). Moreover, stronger activation of the anterior cingulate cortex (ACC) and insula was observed in experimental pain conditions.
Cognitive attention to, or distraction from, pain are important in pain modulation (22–26) in particular to shift the subject’s attention away from clinical pain (27, 28) and, accordingly, reducing its intensity. On a neurobiological level, pain modulation correlates with a profound interaction between central pain-encoding and pain-inhibiting areas (24, 26, 29). In particular, the frontal lobe, the anterior cingulate cortex, the hypothalamus, the insula and the brainstem (nucleus cuneiformis (NCF), and the periaqueductal grey (PAG)) are involved in the descending modulation of pain (30).
Distraction not only leads to a reduction in the intensity of pain (31, 32), but also of itch (33). The neurobiological background of this phenomenon in itch is unknown. Mochizuki et al. (16) showed that itch can be reduced by painful stimuli, leading to an activation of the PAG. In contrast, Papoiu et al. (34) postulated that itch modulation might be contrary to mechanisms known to suppress pain. They found an activation of the reward system (namely the ventral tegmentum) that led to a deactivation of the PAG. A few studies have examined the effect of drugs on itch modulation. Naltrexone (35) reduced fMRI activity during itch perception induced by histamine or capsaicin, while butorphanol suppressed only histamine-induced itch. In the latter condition, structures of the mesolimbic circuit seemed to play an important role during itch suppression (36). As the PAG seems to play not only an important role in pain modulation (by distraction), but also in itch modulation, we examined the activity of the PAG and the NCF during itch with and without distraction. In line with results from the pain field, we hypothesized that comparable activation of brainstem areas and a decrease in itch sensation during distraction should occur. This paper provides additional analyses based on imaging data published in 2013 from our working group (37). Here, we focus on the effects of distraction on activity in the PAG and NCF. In our previous paper, sex-specific differences were observed in localization and itch perception (37). Since no sex-differences were found for itch sensation and brain activity during distraction, data for male and female subjects were pooled in this paper.
Study subjects, induction of itch sensation, the experimental and imaging protocol, and the psychophysical measurements have been described in detail previously (37). The following sections give a short summary.
A total of 33 healthy subjects (age 25.5 ± 3.6 years (mean ± standard deviation; SD)) with no history of chronic disease, allergy or mental illness were included in the study. All subjects gave written informed consent. The study was approved by the ethics committee of the Medical Faculty of Münster, Germany, and was conducted in accordance with the principles of the Declaration of Helsinki.
Itch was induced by applying histamine intradermally via 4 microdialysis fibres (2 in the left forearm and 2 in the left lower leg). Itch stimulation during fMRI scanning was induced by 0.5 ml of 10–4 M sterile histamine solution (Sigma, Deisenhofen, Germany) and terminated by local application of an ice bag and perfusion of 0.7 ml 2% xylocaine-solution (Astra Zeneca, Wedel, Germany).
Magnetic resonance imaging was performed with a 3T-scanner (Gyroscan, Philips, Best, The Netherlands) using a standard receiver head coil. For each subject, 844 echo-planar volumes (EPI) were obtained with the following parameters: TR = 2.5 s, TE = 35 ms, flip angle 90°, matrix dimensions 64 × 64 field of view (FOV) = 210 mm, 36 oblique slices parallel to the AC-PC line; slice thickness: 3.6 mm, pixel size: 3.6×3.6×3.6 mm.
During continuous fMRI scanning the “baseline” condition (50 s with no sensory stimulation) was followed by the histamine perfusion. After 60 s of perfusion a 90 s period of “itch” was initiated, followed by the “Stroop” condition (90 s). During this distraction condition the subjects had to perform a pseudorandomized colour Stroop task (38). During the “itch” condition the subjects looked at a black cross projected onto the screen. At the end of the experiment, the itch sensation was terminated by local anaesthetics and an ice bag (see above). To avoid habituation, the order of the runs (first “itch” or first “Stroop”) and the localization (lower leg or forearm) were altered. There were 4 runs in total.
The identical experiment was repeated with 10 of the 33 volunteers (5 females, 5 males) of the previous experiment approximately 24 months after the first session. In this control condition histamine and xylocaine were replaced by physiological saline solution (0.9% NaCl solution).
Subjects were asked to rate their itch intensity, desire to scratch and pain intensity after the “baseline”, “itch” and “Stroop” condition via a numeric rating scale (NRS) ranging from 0 (no itch/no desire to scratch/no pain) to 10 (most intense itch/desire to scratch/pain imaginable) by using a computer mouse held in their right hand. The NRS values under “itch” and “Stroop” condition were calculated as the mean value of each run.
In addition, immediately before and after the experiment, subjects gave information about their mood and their arousal via a self-assessment manikin (SAM) (39).
Statistical calculations were performed with PASW 21.0 (SPSS Inc., Chicago, IL, USA). Corresponding to the fMRI data a 3-factorial analysis of variance (ANOVA) was performed with the factors order (“first” or “second” condition in the stimulation sequence, 2 levels), condition (“itch”, “Stroop”, 2 levels) and localization (“forearm”, “lower leg”, 2 levels). Because of our initial hypothesis of itch reduction by distraction during the Stroop task we focused on the main effect of condition and the interaction of condition×localization or condition×order.
Furthermore, we used t-tests for dependent variables to measure pre- and post-differences of mood and arousal and differences in reaction time and a non-parametric Spearman’s Rho test to calculate correlations between brainstem activity levels and itch intensity during histamine perfusion.
Functional images were analysed using the general linear model (40) for block designs in SPM8 (Welcome Department of Imaging Neuroscience; London, UK; www.fil.ion.ucl.ac.uk/spm). All images were realigned, normalized to an EPI template (resulting voxel size of 2 mm), spatially smoothed (8 mm FWHM kernel), and high-pass filtered (128 s).
For each subject, data were gathered under 3 conditions: “baseline” condition (20 scans), “itch” condition (36 scans), “Stroop” condition (36 scans) for all runs, resulting in 12 conditions in total (3 conditions×2 runs×2 localizations). Realignment parameters were integrated as regressors into the model. Contrasts were defined as follows: “Stroop”=“Stroop”>“baseline” and “itch”=“itch”>“baseline”. For each subject, 1 BOLD-contrast for each stimulation at each extremity was determined as a function of BOLD-signal changes, so there were 4 BOLD-contrasts in total: first stimulation forearm (A1_itch; A1_Stroop), first stimulation lower leg (B1_itch; B1_Stroop), second stimulation forearm (A2_itch; A2_Stroop) and second stimulation lower leg (B2_itch; B2_Stroop).
The individual BOLD-contrasts were transferred into a 3 factorial ANOVA (factors order (“first” or “second” position in the stimulation sequence, 2 levels), condition (“Stroop”, “itch”, 2 levels) and localization (“forearm”, “lower leg”, 2 levels)) using SPM8 “full factorial” design. The main effect of condition (and its differential contrast of each condition) and its interactions with order and localization were assessed. Family-wise error correction (p < 0.05) with a minimum cluster size of 20 voxels were defined as levels of significance.
BOLD-Signal in brainstem (PAG, NCF). Because of our initial hypothesis that itch inhibition correlates with brainstem activation during distraction (Stroop task) we determined the condition-dependent course of significant brainstem activations (eigenvariate). The region of interest was chosen according to the analysis of Irene Tracy’s group (41, 42). During whole-brain analysis we found significant brainstem activation (x,y,z-coordinate -8 -26 -10) and the eigenvariates (sphere of 5 mm) of the left (-8 -26 -10) and right (contralateral, 8 -26 -10) cluster were extracted in SPM8. To control whether this activation pattern was based on a pure Stroop-effect (and not itch-related) the eigenvariate of the cluster activation of the 10 participants during our control experiments (saline perfusion) was also evaluated.
“Stroop”condition. After deleting inconsistent results (participants pushed the button too late or too early), 4,582 tasks were included in the analysis with 4,158 correct answers. The reaction time of the correct answers decreased over the course of the experiment due to a practice effect (A1 = 1,448.9 ± 302.3, n = 993; B1 = 1,362.0 ± 276.0, n = 1,042; A2 = 1,364.2 ± 302.1, n = 1,040; B2 = 1,316.3 ± 274.1, n = 1,083). The low drop-out rate of approximately 3% and an error rate of approximately 10% indicate that subjects performed the task as expected and were distracted. A detailed analysis is given in our previous paper (37).
Valency and arousal. There was no significant difference in the extent of arousal at the beginning and at the end of the experiment (0.8 ± 1.0, 0.9 ± 0.7; p = n.s.). In contrast, at the end of the experiment, the mood of subjects was significantly more negative than at the beginning (2.5 ± 0.9, 2.8 ± 0.8; p = 0.049).
There were no significant differences in arousal (0.7 ± 0.7, 0.6 ± 0.7; p = n.s.) or valency (2.9 ± 0.7, 2.6 ± 1.0; p = n.s.) at the beginning and the end of the experiment during the control experiment.
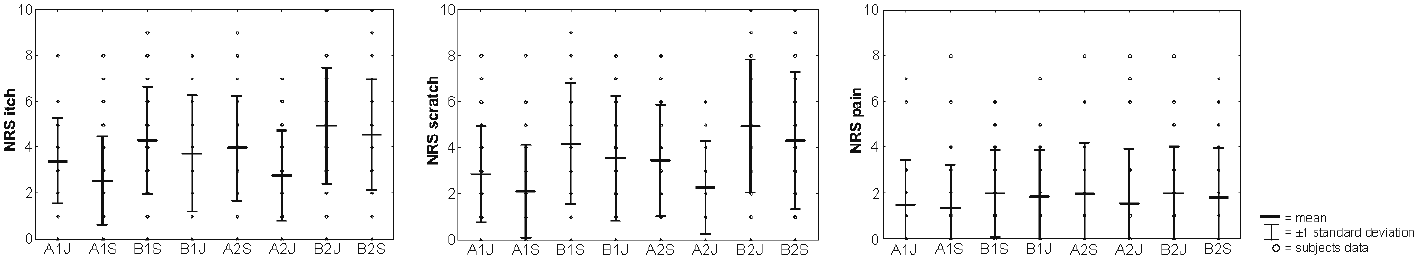
Itch sensation, desire to scratch and pain. The participants had a mean itch intensity of 3.77 (min–max: 1–8), a mean desire to scratch of 3.46 (min–max: 0–8) and a mean pain intensity of 1.72 (min–max: 0–7). For further details see Fig. 1.
Fig. 1. Mean intensities and standard deviations of psychophysical data (numeric rating scale; NRS) of itch intensity, desire to scratch and pain during each run. ?A1J = first “itch” condition at the forearm; A1S = first “Stroop” condition at the forearm; B1S = first “Stroop” condition at the lower leg; B1J = first “itch” condition at the lower leg; A2S = second “Stroop” condition at the forearm; A2J = second “itch” condition at the forearm; B2J = second “Stroop” condition at the lower leg; B2S = second “Stroop” condition at the lower leg.
In contrast to our hypothesis, the 3-factorial ANOVA revealed that the Stroop task did not reduce itch intensity (F-score = 0.61, p = 0.44), but localization (F-score = 17.12, p < 0.001) and order (F-score = 29.30, p < 0.001) showed significant main effects on itch intensity. The interaction condition×order (F-score = 0.50, p = 0.49) and condition×localization (F-score = 0.16, p = 0.69) revealed no significant effects. Similarly, the Stoop task did not significantly reduce the desire to scratch (F-score = 0.49, p = 0.49).
In contrast, the factors localization (F-score = 24.70, p < 0.001) and order (F-score = 23.50, p < 0.001) showed significant main effects. The interactions condition×order (F-score = 0.72, p = 0.40) or condition×localization (F-score = 0.57, p = 0.46) were not significant. Concerning pain intensity there was neither a significant effect for condition (F-score = 0.47, p = 0.50) nor localization (F-score = 2.75, p = 0.11) but for order (F-score = 5.6, p = 0.024). The interactions condition×order (F-score = 1.01, p = 0.32) and condition×localization (F-score = 0.51, p = 0.48) were not significant.
During saline perfusion, the participants had a mean itch intensity over all conditions of 0.36 (min–max: 0–4), a mean of desire to scratch of 0.23 (min–max: 0–3) and a mean pain intensity of 0.88 (min–max: 0–8). This was significantly different from the histamine perfusion for itch intensity and desire to scratch (p ≤ 0.001), but not for pain intensity (p = n.s.). In a 3-factorial ANOVA with order, localization and condition, we found a main effect of order (F-score = 7.3, p = 0.024) for itch sensation, but no main effect for localization and no significant effects for the desire to scratch. Because no itch sensation was experienced in the saline condition no further statistical analysis was performed.
Three-factorial ANOVA. There was widespread activation of the occipital gyrus, certainly due to the visual stimuli during the main effect of condition. Furthermore, there was activation of both frontal gyri (including dorsolateral prefrontal cortex (DLPFC) and pre-supplementary motor area, BA 6, 9, 46), left thalamus, right insula, left brainstem, both lentiform nuclei, left middle temporal gyrus (BA 39) and right parahippocampal gyrus (BA 27) and left precuneus (BA 31) (Table SI).
Differential contrast analyses (t-tests) revealed that these brain activations were mostly driven by the “Stroop” condition (“Stroop”>“itch”) and, most importantly, showed strong brainstem activation during “Stroop”. The reverse contrast (“itch”>“Stroop”) presented activation of the right insula, left precuneus (BA 31), left middle temporal gyrus (BA 39) and right posterior cingulate gyrus (BA 31) and no brainstem activation (Table I, Fig. 2).
Table I. Differential contrasts of “itch” and “Stroop” condition of the 3-factorial analysis of variance (ANOVA) during histamine perfusion
Table I. Differential contrasts of “itch” and “Stroop” condition of the 3-factorial analysis of variance (ANOVA) during histamine perfusion
Fig. 2. Significant functional magnetic resonance imaging (fMRI) signal intensity changes for the differential contrasts of “Stroop”>“itch” (red) and “itch”>“Stroop” (yellow) in the 3-factorial analysis of variance (ANOVA) with the factors order, condition and localization. *The results are family-wise error-corrected (p < 0.05, voxel threshold > 20 voxels).
The interactions condition×order and condition× localization did not present any suprathreshold clusters.
During the control experiment with saline perfusion the main effect of condition revealed activation of the left cerebellum and occipital brain structures (BA 17, 18) as well as right parietal gyrus, right precuneus, left middle frontal gyrus, including DLPFC (BA 9, 46) and left inferior parietal lobule (BA 40) (Table SII).
Concerning the differential contrasts (t-tests) the contrast “Stroop”>“itch” showed activation of the left cerebellum, left lingual gyrus (BA 17), left cuneus (BA 18), right occipital gyrus (BA 18) and the right superior and left inferior parietal gyrus (BA 7, 40), right precuneus (BA 7) and left inferior and middle frontal gyrus (BA 7, 9) (Table II). The differential contrast of “itch”>“Stroop” did not reveal any suprathreshold clusters.
Table II. Differential contrast of “itch” and “Stroop” condition of the 3-factorial analysis of variance (ANOVA) during saline perfusion
The interaction condition×order, as well as the interaction condition×localization, did not reveal any supra-threshold clusters.
BOLD brainstem activity. The eigenvariate as a measure of activation indicates higher activations of the left brainstem cluster during “Stroop” condition compared with the “itch” condition (Fig. 3). Statistical comparison of activation intensities in every single run revealed significant elevated intensities for those runs in which the “itch” condition was first in the sequence (first-run forearm: T-score = –5.67, p-score < 0.001, second-run lower leg: T-score = –3.22, p-score = 0.003). For the other 2 runs, where the Stroop task was first, we could not find any significant differences in brainstem activation pattern (second-run forearm: T-score = –1.10, p-score = 0.28, first-run lower leg: T-score = –0.69, p-score = 0.49).
Fig. 3. Eigenvariates of brainstem activation during “itch” and “Stroop” during histamine perfusion. A significant difference between these 2 conditions can be found only during the first stimulation at the forearm and the second stimulation at the lower leg in the left brainstem. ?*p < 0.05, **p < 0.01, ***p < 0.001. ?A1J = first “itch” condition at the forearm; A1S = first “Stroop” condition at the forearm; B1J = first “itch” condition at the lower leg; B1S = first “Stroop” condition at the lower leg; A2J=second “itch” condition at the forearm; A2S = second “Stroop” condition at the forearm; B2J = second “Stroop” condition at the lower leg; B2S = second “Stroop” condition at the lower leg.
Moreover, the left brainstem activity correlated significantly with itch intensity (r = –0.13, p = 0.035), but not with right brainstem activity ( r =–0.03, p = 0.58).
During saline perfusion without appreciable itch sensation, there was no consistent brainstem activation during the Stroop task (Fig. 4).
Fig. 4. Eigenvariates of brainstem activation during “itch” and “Stroop” during saline perfusion. A significant difference between these 2 conditions can be found only during the second stimulation at the forearm. ?*p < 0.05. ?A1J = first “itch” condition at the forearm; A1S = first “Stroop” condition at the forearm; B1J = first “itch” condition at the lower leg; B1S = first “Stroop” condition at the lower leg; A2J = second “itch” condition at the forearm; A2S = second “Stroop” condition at the forearm; B2J = second “Stroop” condition at the lower leg; B2S = second “Stroop” condition at the lower leg.
The present study is a new analysis of data obtained in a former study (37) in which sex-specific differences in central itch perception and modulation were analysed. In the present study, the effect of distraction by a colour Stroop task on cerebral itch processing was investigated. In contrast to our initial hypothesis, the Stroop task did not reduce any itch-related sensation (intensity, desire to scratch, pain). However, distraction activated, in particular, the DLPFC, brainstem and thalamus, all structures well known to be involved in pain modulation.
The analgesic effect of distraction is a well-known phenomenon in experimental (32) and clinical settings (31). Distraction also reduces clinical itch, as shown during experimental distraction in a virtual reality and audiovisual distraction environment (33).
In contrast to the study by Leibovici et al. (33), we did not find distraction to reduce itch intensity or urge to scratch. The difference might be related to the selection of our participants (younger and healthy subjects), different itch stimulation (experimental vs. clinical) and lower itch intensity in our study. Moreover, Leibovici’s distraction via virtual reality might have been a more effective strategy to reduce itch intensity compared with the Stroop task.
Analgesic effects of distraction (including Stroop-induced) correlate with increased PAG and NCF activation (26, 30, 42, 43) which emphasizes the importance of brainstem in pain reduction. In our study we found Stroop-related activation of the brainstem with increased activation intensity (eigenvariate) of the left ipsilateral NCF during “Stroop” condition of the 1st and 4th runs (see Fig. 3).
Mochizuki et al. could show that a reduction in experimental itch sensation through an additional painful cold stimulus leads to PAG activation, comparable to diffuse noxious inhibitory control (DNIC) in pain (16). Therefore, the same brainstem areas seem to play a role in pain and itch reduction. The important role is further supported by the observed negative correlation of itch intensity with brainstem activation.
Interestingly, Papoiu et al. (34) hypothesized that itch scratch-evoked reward is linked to deactivation of the PAG. This deactivation was associated with activation of the reward system, especially the ventral tegmentum. Thus, PAG modulation appears to differ between pain-induced reduction in itch (either via a painfully cold stimulus or via distraction) and scratch-induced pleasure.
Moreover, Vierow et al. (35) and Papoiu et al. (36) could reduce itch sensation by naltrexone and butorphanol. While naltrexone reduced the fMRI activity during itch perception induced by histamine and capsaicin, butorphanol led to a reduction in histamine-induced itch only and perfusion changes in the midbrain, thalamus, S1, insula, and cerebellum. During butorphanol infusion, the ventral tegmental area (VTA), periaqueductal gray (PAG), raphé nucleus, thalamus, precuneus and cerebellum were activated. As the PAG is activated in our study, too, it might be feasible to assume that cognitive distraction and drugs lead to corresponding activations of inhibiting systems.
A Stroop task as an instrument of focused attention leads to activation of the anterior cingulate cortex, dorsolateral prefrontal cortex and/or parietal cortex (44–52). However, we assume that the increased DLPFC activation does not merely reflect the effect of attention, but is also involved in pain and itch modulation. Pain studies report that the DLPFC is responsible for anticipation of pain and emotional weighting of pain (53). In itch studies, the DLPFC (with premotor and parietal areas) may reflect the planning of the motor response to itch (scratching) or the attenuation of the desire to scratch (12, 17). Increased DLPFC activity during repetitive scratching has also been reported, suggesting a link with scratch-induced inhibition of itch and induction of pleasure and reward (12, 54). Thus, activation of DLPFC might also be directly linked to modulation of the itch sensation. Furthermore, the DLPFC interacts with the descending pain modulation system, particularly PAG and NCF (43, 55), representing another possible link to itch modulation. These results are supported by our control experiment, in which DLPFC activation was unilateral during “Stroop”>“itch” and not bilateral, as during the histamine experiment.
The activation of thalamus during itch sensation is well known (11, 13). Schneider et al. (20) found a significantly higher activation of an orbito-frontal-pallidum-thalamus circuit for patients with atopic dermatitis, and hypothesized that this circuit could facilitate scratching behaviour. Thus, compared with our study the lentiform nucleus might play a role in modulation of itch and scratching desire by distraction.
First, the intensity of itch sensation might not have been sufficient to distinguish clearly between both conditions (“itch”, “Stroop”) and to decide whether the itch sensation increases or decreases noticeably. As the whole experiment took more than 30 min we cannot rule out that fatigue of our subjects influenced the results.
Secondly, we do not have a “pure” itch sensation, but a certain mixture with pain sensation. The pain sensation might be provoked by the histamine solution, which has a specific “burning” effect. However, the pain sensation is much lower than the itch sensation, so we assume that pain sensation plays only a minor role. Moreover, the low drop-out rate of 2.9% and an error rate of 9.8% indicate that subjects performed the task as expected and were distracted.
Finally, we did not perform a single study analysis on brainstem level. We cannot rule out that our minimum cluster size was too large and PAG brain activity measurements may have been influenced by signals from neighbouring structures. Furthermore, we did not perform a separate normalization procedure of the brainstem, besides the normalization in SPM in accordance with the MNI template during pre-processing, which might have influenced the localization of our brainstem cluster.
In conclusion, distraction paradigms have been shown to reduce itch and pain via activation of overlapping brain regions, especially by the brainstem. However, our data suggest that even a clear activation of DLPFC, brainstem and thalamus by a Stroop task is not sufficient to reduce histamine-induced itch sensation.
The authors would like to thank Hermann Schillers, Department of Physiology, University of Münster for technical support, and Stefanie Schelzig and Özkan Genc, Department of Clinical Radiology, University of Münster for fMRI measurements.
The authors acknowledge support from the Open Access Publication Fund of the University of Münster.
The authors declare no conflicts of interest.


 Click to show fullsize
Click to show fullsize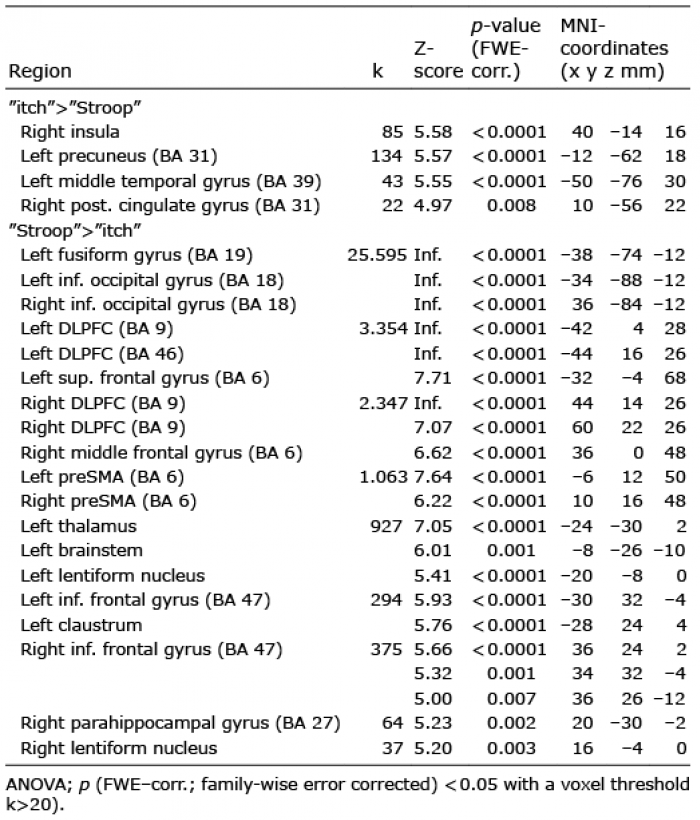 Click to show fullsize
Click to show fullsize Click to show fullsize
Click to show fullsize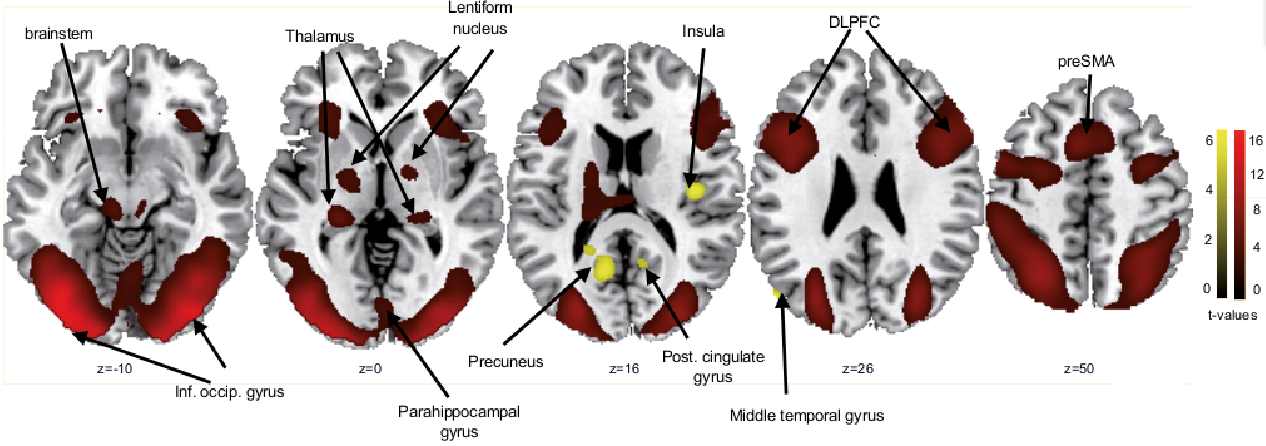 Click to show fullsize
Click to show fullsize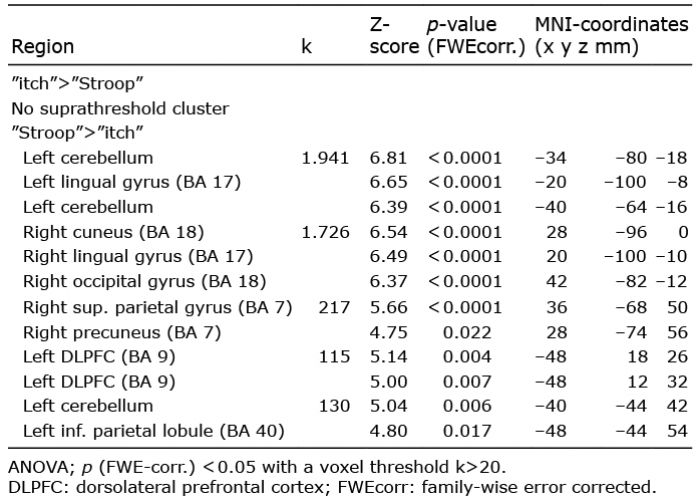 Click to show fullsize
Click to show fullsize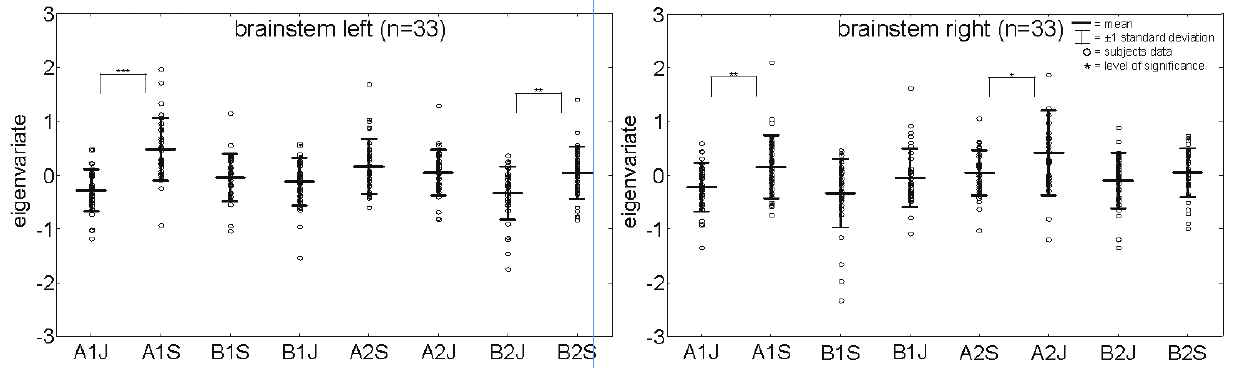 Click to show fullsize
Click to show fullsize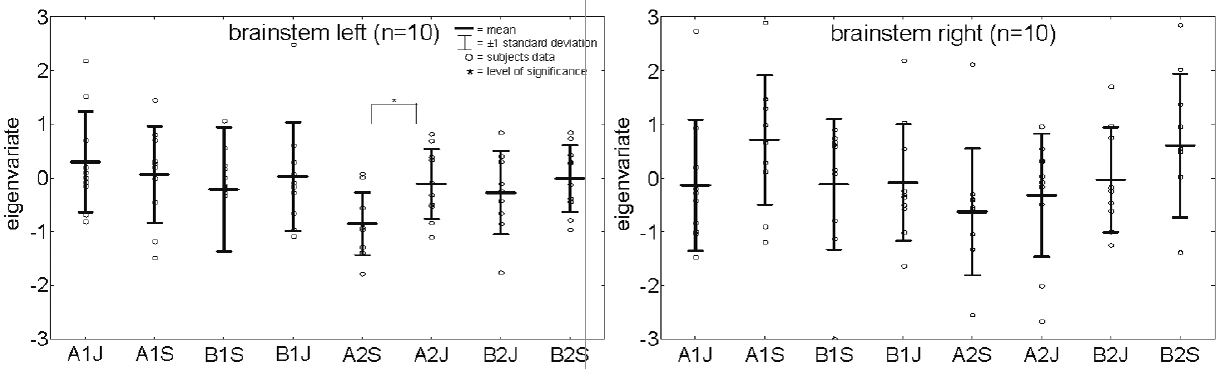 Click to show fullsize
Click to show fullsize