


1Biofilm Research Laboratory, Institute of Dental Sciences, Faculty of Dental Medicine, The Hebrew University of Jerusalem, Jerusalem, 2The Cutaneous Microbiology Laboratory, The Skin Research Institute, The Dead Sea and Arava Science Center, Masada, 3Department of Mathematics and Computer Science, The Open University of Israel, Raanana, 4Department of Physics of Complex Systems and 7Department of Molecular Cell Biology, Weizmann Institute of Science, Rehovot, 5Department of Dermatology, Rabin Medical Center, Beilinson Hospital, Petach Tikva, and 6DMZ Medical Center, M. P. Dead Sea Ein Bokek, Israel
#These authors contributed equally to this work.
Dead Sea climatotherapy (DSC) is a therapeutic modality for a variety of chronic skin conditions, yet there has been scarce research on the relationship be-tween the cutaneous microbiota and disease states in response to DSC. We characterized the skin bacterial and fungal microbiome of healthy volunteers who underwent DSC. Bacterial community diversity remained similar before and after treatment, while fungal diversity was significantly reduced as a result of the treatment. Individuals showed greater inter-individual than temporal bacterial community variance, yet the opposite was true for fungal community composition. We further identified Malassezia as the genus driving temporal mycobiome variations. The results indicate that the microbiome remains stable throughout DSC, while the mycobiome undergoes dramatic community changes. The results of this study will serve as an important baseline for future investigations of microbiome and mycobiome temporal phenomena in diseased states.
Key words: skin microbiome; skin mycobiome; climatotherapy.
Accepted Aug 16, 2017; Epub ahead of print Aug 17, 2017
Acta Derm Venereol 2018; 98: xx–xx.
Corr: Shiri Meshner, The Cutaneous Microbiology Laboratory, The Skin Research Institute, The Dead Sea and Arava Science Center, Masada 8693500, Israel. E-mail: shiri.meshner@evogene.com
The introduction of 16S amplicon sequencing, followed by whole-genome shotgun sequencing, has added new layers of understanding to our knowledge of skin commensals and pathogens. Whereas site-specificity of microorganisms on skin had been known for decades (1), skin microbiome studies have enabled a clearer elucidation of skin habitats and their resident bacteria (2, 3). In addition, the degree of temporal stability of the human skin microbiome has been expounded through both 16S and metagenomics studies (4, 5). With this knowledge in hand, many groups have moved on to elucidate cutaneous microbial community perturbations in a myriad of disease states, including atopic dermatitis (6, 7), acne vulgaris (8, 9), psoriasis (10), vitiligo (11) and Leishmaniasis (12).
Recent studies on skin-resident microbiota have expanded to include fungal species. Importantly, community differentiation based on biogeographical sites is less pronounced in fungi than in bacteria, with only species-level classification allowing for such distinctions (13). Metagenomic studies have shown that the mycobiome is stable over time, much like the microbiome (5), while amplicon-based studies, usually acquired by amplifying the internal transcribed spacer region (ITS) of the ribosome, have shown that community diversity converges from childhood to adulthood (14). To the best of our knowledge, neither method has been used robustly to explore the connection between skin disease states and the mycobiome.
Dead Sea climatotherapy (DSC), or the combined effect of milder ultraviolet A (UVA) and ultraviolet B (UVB) radiation, Dead Sea baths, atmospheric bromide and magnesium, and relaxation has been shown to be an effective treatment for several skin diseases (15). Benefits of DSC over alternative treatment methods include longer remission periods, rare side-effects, and psychological improvement (16–18). In addition, DSC can be a more economic treatment method than expensive biologicals (17, 19). Even.-Paz et al. (20) showed that the main therapeutic element of DSC is sun exposure, with Dead Sea water baths potentially enhancing the effects of the sun. Subsequently, Kudish et al. (21) showed that UVB solar radiation is attenuated in the Dead Sea basin, such that “incident solar UVB has a higher ratio of therapeutic to erythema radiation relative to other sites”. We hypothesized that clinical improvements in patients undergoing DSC are accompanied by microbial community shifts similar to those observed with other treatment methods. However, before examining the skin microbiome of diseased individuals in response to DSC, we aimed to characterize the degree of stability of the skin bacterial and fungal microbiome of healthy individuals, in an attempt to isolate the effect of the clinical therapeutic element from the environment.
This study was approved by the Institutional Review Board (Helsinki Committee) of the Rabin Medical Center, Petah Tikva, Israel (approval number 0153-12-RMC). Eleven adult guardians accompanied a group of adolescent patients undergoing DSC in March 2015 and were invited to enroll in the study. They provided written informed consent. Volunteers were sampled at the commencement of treatment at the DMZ Medical Center (Ein Bokek, Israel) and once again following a complete course of DSC. DSC entails a thorough medical examination at baseline, at which point the climatotherapy specialist (a dermatologist) prescribes a detailed schedule of sun exposure and Dead Sea bathing for the 21-day period. The specialist takes into account skin type, seasonal fluctuations in ultraviolet (UV) radiation and, where relevant, disease severity (22). None of the volunteers were pregnant at the time of sampling, none reported a personal history of skin diseases, and none had used antibiotics in the month prior to sampling.
Volunteers were asked not to wash or apply cosmetics or cream on the antecubital fossa (inner elbow) for 12 h prior to sampling. The sex and age of participants were recorded. Samples were taken from a 2 × 2 cm area of the antecubital fossa using a sterile rayon-tipped swab (COPAN, Brescia, Italy) soaked in sterile 0.15 M NaCl with 0.1% Tween 20 (J.T. Baker) before and after DSC (10). Swabs were immediately transferred to –80°C and stored until DNA extraction.
All pre-PCR, post-clinical handling and experimental procedures were carried out in a DNA/RNA UV Cleaner-Recirculator (Biosan, Latvia) located in a PCR-free facility. DNA was extracted using the MoBio PowerSoil DNA Extraction kit following the manufacturer’s protocol and supplementary instructions for low bio-burden samples. A multiplex PCR primer-set was used to simultaneously amplify 5 variable regions of the 16S rRNA gene. For ITS amplification, the 18SF (GTAAAAGTCGTAACAAGGTTTC) and 5.81R (GTTCAAAGAYTCGATGATTCAC) primer pair was used (13), with the addition of Illumina overhang adapter sequences. PCR conditions were as follows: 98°C for 2 min, followed by 35 cycles of 98°C for 10 s, 61°C for 15 s (51°C for ITS) and 72°C for 35 s, and a final extension step of 72°C for 5 min. PCR reaction mix contained 10 µl Kapa HiFi HotStart Readymix (Kapa Biosystems, Wilmington, MA, USA), 0.4 µl primer mix (10 µM of each primer), 7.6 µl Molecular Biology Water (Sigma, St Louis, MO, USA), and 2 µl template DNA. PCR was set up in triplicate. Following PCR, sample triplicates were pooled and amplicons were cleaned using Agencourt AMPure XP kit (Beckman Coulter, Indianapolis, USA), and barcodes and adapters were added in a second PCR reaction. Subsequently, samples were pooled and cleaned using the Agencourt AMPure XP kit (Beckman Coulter). Negative control reaction blanks were included for the extraction and PCR steps, and were run on a 2% agarose gel, whereby the lack of a visible band confirmed proper and clean implementation of the protocol. Sequencing was carried out at Hebrew University’s Genomic Applications Laboratory (Ein Kerem, Jerusalem, Israel) on an Illumina MiSeq instrument, using the v2 2×150 kit (16s) or the v2 2×250 kit (ITS).
Demultiplexed 16S rRNA reads were passed to the SMURF software package (source code available at https://github.com/NoamShental/SMURF), an updated version of the COMPASS software package, which allows for computational reconstruction of the 16S rRNA gene using sequence reads from any number of variable regions of the gene. SMURF provides a joint estimate of the microbial community composition, enabling both accurate and extremely high resolution microbial profiling (23). Demultiplexed ITS reads were processed with QIIME and closed reference Operational Taxonomic Units (OTUs) were picked using the Findley ITS database (13, 24).
The R statistical software package was used for all statistical tests (25). Relative abundance of phyla and genera are presented as the mean ± SEM. Taxonomy tables (16S) and OTU tables (ITS) were input into the phyloseq package in R to obtain alpha and beta diversity metrics. The Shannon diversity index and Bray-Curtis distance were used as measures of alpha and beta diversity, respectively. The Wilcoxon signed-rank sum test was used to determine significance of temporal changes in alpha diversity, phyla and genera. All p-values were adjusted according to the false discovery rate method. Permutational multivariate analysis of variance (PERMANOVA) was used to determine significance in beta diversity changes and Pearson’s product moment correlation was used to determine correlations between variables.
Twenty-two skin samples from 11 adult volunteers were obtained for this study. Following quality filtering, a total of 1.7 million 16S high-quality sequences were input into the SMURF algorithm. Samples were subsequently rarefied to 3,600 reads per sample, resulting in the loss of one sample and the subsequent exclusion of its paired sample. For the ITS sequences, we obtained 340,000 high-quality reads for 18 samples and subsequently rarefied to 4,000 reads per sample. As a result of rarefaction, one volunteer’s samples were excluded from the study.
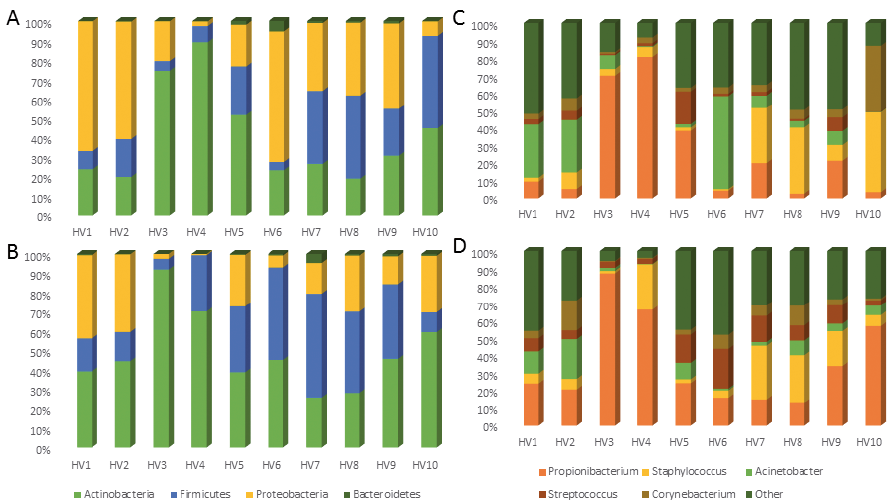
At the phylum level, samples were dominated by Actinobacteria (baseline 40.5 ± 7.7%, post-DSC 48.9 ± 6.3%), Firmicutes (baseline 22.2 ± 5%, post-DSC 29.2 ± 5.2%), Proteobacteria (baseline 36.0 ± 7.4%, post-DSC 20.7 ± 4.7%), and to a lesser extent Bacteroidetes (baseline 1.1 ± 0.5%, post-DSC 0.9 ± 0.4%), both before and after DSC (Fig. 1a, b). The dominance of the aforementioned 4 phyla in the skin microbiome has been documented extensively previously (26, 27). Slight outgrowth of Actinobacteria and Firmicutes at the expense of Proteobacteria was recorded, yet it was not statistically significant (p > 0.1). Therefore, none of the phyla were significantly altered between sampling sessions, leading us to conclude that the phylum-level microbiome of healthy individuals is stable throughout treatment at the Dead Sea.
Fig. 1. Relative abundances of (A, B) bacterial phyla and (C, D) genera of healthy volunteers (A, C) prior to and (B, D) after Dead Sea climatotherapy.
At the genus level, 5 genera accounted for 68% of the bacterial community. Consistent with previous reports on the skin microbiome (3), specifically that on the antecubital fossa, our samples were dominated by Propionibacterium (baseline 25.6 ± 9.0%, post-DSC 35.8 ± 8.1%), Staphylococcus (baseline 14.7 ± 5.3%, post-DSC 12.9 ± 3.7%), Acinetobacter (baseline 14.3 ± 5.6%, post-DSC 6.8 ± 2.2%), Streptococcus (baseline 4.3 ± 1.8%, post-DSC 9.5 ± 2.1%) and Corynebacterium (baseline 7.2 ± 3.4%, post-DSC 5.5 ± 1.7%) (Fig. 1c, d). While slight variations in the relative abundance of each genera are noted, none are statistically significant (p > 0.1).
To assess the temporal stability of the skin microbiome, we first calculated the Shannon diversity index for each sample and its coefficient of variance (CV). Low CV values indicate community stability, while high CV values indicate higher community variability (4). Community Shannon diversity values were nearly indistinguishable between time-points, and low CV values indicated almost universal (9/10) temporal stability of alpha diversity in our cohort (Fig. 2a, c). There was no statistical significance between the baseline and post-DSC samples when testing for change in community diversity as measured by the Shannon index. We then calculated the Bray-Curtis distances between all samples and explored whether community composition changed as a result of DSC using PERMANOVA. Community composition was significantly different between individuals (p < 0.05), but not between time-points (p = 0.08) (Fig. 3a). Taken together, our results indicate that the temporal stability of the human skin antecubital fossa microbiome is not affected by DSC. We do, however, note that 16S community analysis does not distinguish between living, dormant and dead members of the microbial community. Previous studies have reported that phototherapy can affect skin commensal physiological states, including the production of toxins and super-antigens (28, 29). Therefore, the observed stability of the healthy human skin microbiome following DSC reflects only total community membership, and does not take into account molecular and biochemical capabilities. In addition, several culture-based studies have indicated that skin commensals are halotolerant, and therefore the microbial community may not be affected dramatically by exposure to the high salinity of the Dead Sea (30, 31).
Fig. 2. Shannon diversity boxplots of both (A) bacterial and (B) fungal communities before and after treatment, as well as the coefficient of variance measure of community stability for (C) both bacteria and fungi and (D) inverse correlation between Malassezia relative abundance and Shannon diversity of the mycobiome.
Fig. 3. Principal coordinates analysis (PCoA) plots based on Bray-Curtis distances of (A) bacterial and (B) fungal microbiomes coloured by time-point.
In addition to profiling the resident bacterial community, we also characterized the cutaneous fungal community in 8 volunteers. More than 99% of the mycobiome comes from the 2 phyla Ascomycota (baseline 58.3 ± 10.6%, post-DSC 30.0 ± 10.4%) and Basidiomycota (baseline 41.3 ± 10.5%, post-DSC 69.5 ± 10.4%), consistent with previous reports of the skin mycobiome (13). Both Basidiomycota and Ascomycota change significantly as a result of DSC (p < 0.05) (Fig. 4).
Fig. 4. Relative abundances of (A, B) fungal phyla and (C, D) genera of healthy volunteers (A, C) prior to and (B, D) after Dead Sea climatotherapy.
At the genus level, we identified 12 genera that accounted for 93% of fungal reads. Critically, the genus Malassezia, the dominant member of the adult human mycobiome (14) and the only genus present in every sample in our study, undergoes a dramatic shift in its relative abundance (baseline 30.5 ± 10.4%, post-DSC 66.8 ± 11.0%, p < 0.05) following DSC (Fig. 4c, d). Malassezia outgrowth has been associated with various skin diseases including pityriasis versicolor, seborrhoeic dermatitis, and Malassezia folliculitis (32). However, the molecular mechanisms by which Malassezia shift from commensal residents of the skin to pathogens are poorly understood, and therefore Malassezia outgrowth following DSC absent detrimental clinical presentations can indicate a restorative property of DSC on the healthy skin mycobiota. This outgrowth of Malassezia following DSC can explain the overall observed drop in community diversity as measured by the Shannon index (p < 0.05, Fig. 2b, d). In addition, higher overall CV values indicate lower community stability, as evident by the change in community composition following DSC (Fig. 2c). When examining temporal changes through the lens of beta diversity, the majority of post-DSC samples cluster together on a principal coordinates analysis (PCoA) (Fig. 3b). Notable outliers are volunteers 10 and 11, whose post-DSC fungal signature are not dominated by Malassezia (Fig. 4d). Interestingly, these 2 individuals entered the study with the smallest relative amount of Malassezia, potentially indicating a need for a pre-existent core community to support outgrowth of Malassezia throughout treatment. In contrast to the microbiome, fungal community composition was significantly different between time-points (p < 0.05) but not so between individuals (p > 0.1), as identified by PERMANOVA. Taken together, these results indicate that DSC significantly affects the temporal stability of the healthy human skin mycobiome without significantly affecting the human skin microbiome. Due to the dearth of studies examining either the effect of UVB or high salinity on Malassezia, there is little existing knowledge upon which to theorize regarding the mechanism governing the outgrowth of Malassezia following DSC. However, lipophilic Malassezia may take advantage of differing topographical biochemistry induced by DSC, allowing for its increased colonization.
DSC is an efficacious form of therapy for various skin and systemic afflictions. We conducted a small-scale clinical study to measure the effect of DSC on the bacterial and fungal communities that colonize healthy human skin. The results indicate that the healthy human skin microbiome remains undisturbed following DSC, including the prominent genera Staphylococcus, Propionibacterium, and others, while skin fungal communities become less diverse as a result of Malassezia outgrowth during DSC. Malassezia is the dominant fungal genera in healthy adults, yet specific species of Malassezia have been associated with human skin afflictions, while other species are considered harmless skin commensals. Therefore, the clinical importance of the increase in relative abundance of Malassezia reported here necessitates further investigation and species-level characterization of Malassezia. The results reported here highlight the stability of the common bacterial genera comprising the human skin microbiota when exposed to therapeutic dosages of UV radiation. Further studies, incorporating larger cohorts and different clinically relevant topographical sites, should be initiated to further our understanding of the reported observations. The results of this study will serve as an important baseline for future investigations of microbiome and mycobiome temporal phenomena in diseased states, such as atopic dermatitis, psoriasis, vitiligo and chronic fatigue syndrome, following DSC. In addition, DSC may offer a therapeutic modality towards the restoration of a normative level of Malassezia on the skin.
The authors would like to thank the volunteers who participated in this study. In addition, we acknowledge Shira Wax and Ira Mezin for assisting in ethics approval and planning. Dr Miriam Kott, Dr Idit Shiff and Dr Abed Nasereddin of the Hebrew University of Jerusalem Genomic Applications Laboratory provided sequencing support.
This study was approved by the Helsinki Committee of the Rabin Medical Center, Petah Tikva, Israel (Approval Number 0153-12-RMC). Volunteers provided written informed consent prior to participating in the study.
This study was supported by the Israeli Ministry of Science and Technology Grant 3-11174 and by the Norwegian Dead Sea Medical Foundation. Michael Brandwein is a recipient of the Kaete Klausner Fellowship.
Conflict of interest: MH serves as medical director of the DMZ Medical Center. The other authors declare no conflicts of interest.


 Click to show fullsize
Click to show fullsize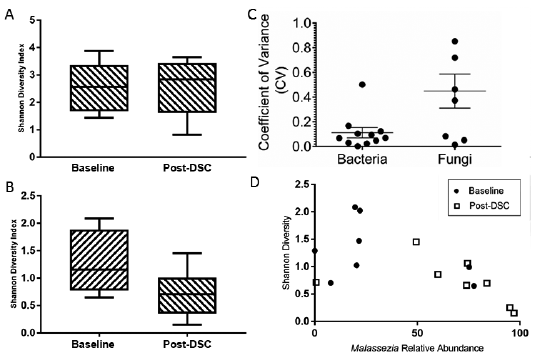 Click to show fullsize
Click to show fullsize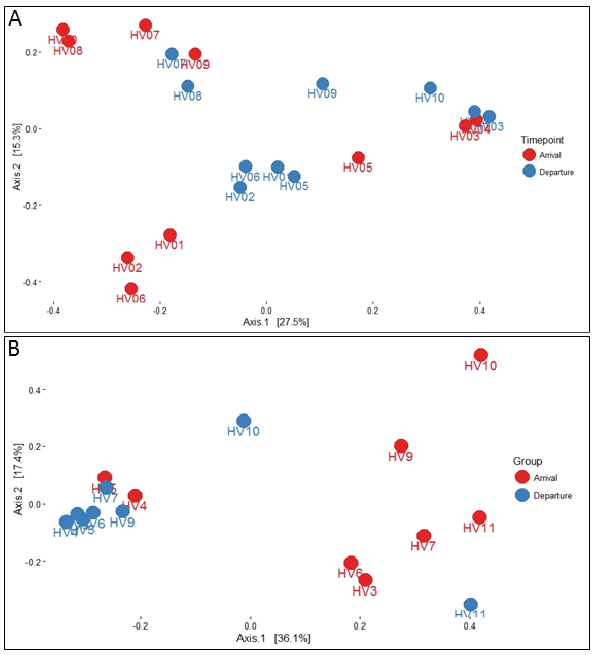 Click to show fullsize
Click to show fullsize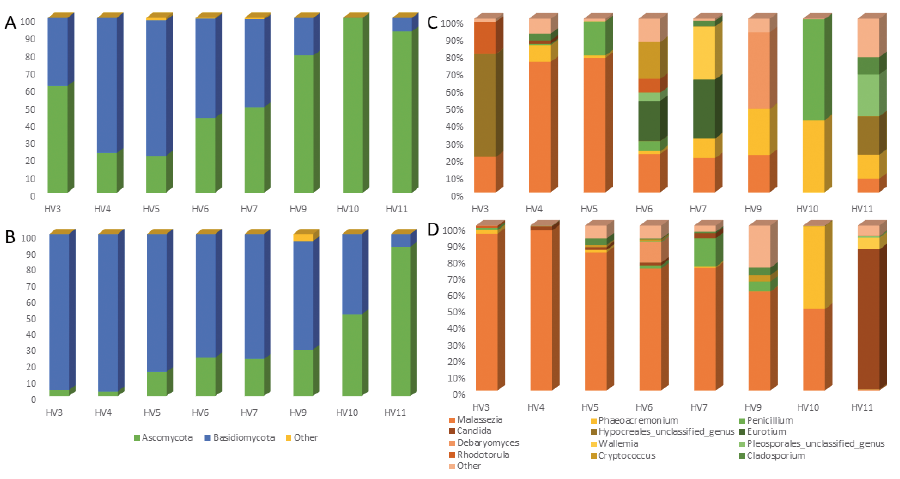 Click to show fullsize
Click to show fullsize