


Department of Dermatology, Nara Medical University School of Medicine, 840 Shijo, Kashihara, Nara 634-8522, Japan. *E-mail: asadah@naramed-u.ac.jp
Accepted Sep 13, 2017; Epub ahead of print Sep 13, 2017
Drug-induced hypersensitivity syndrome (DIHS)/drug reaction with eosinophilia and systemic symptoms (DRESS) is a severe adverse cutaneous drug reaction associated with the reactivation of human herpesvirus 6 (HHV-6). In DIHS, HHV-6 is generally reactivated 2–3 weeks after the onset of a rash, and such reactivation is associated with the flare-up of clinical symptoms (1). The reactivation of HHV-6 usually occurs as a transient event; however, in rare cases HHV-6 DNA continues to be detected long after the onset of the condition, which is sometimes associated with frequent recurrence of clinical symptoms, such as skin rashes. There has been only one report of a case of DIHS involving a persistent HHV-6 infection (2). We report here 3 cases of DIHS in which HHV-6 DNA was detected in the patients’ peripheral blood mononuclear cells (PBMC) long after resolution of their DIHS. We also demonstrated that CD4 T cells were the main contributors to the PBMC HHV-6 DNA load throughout the patients’ clinical courses, while in the early stages of their conditions CD14+ monocytes and other types of PBMC also harboured HHV-6 DNA.
The characteristics of the 3 patients with DIHS are listed in Table SI. Blood samples were obtained from each patient after the onset of a rash. PBMC were isolated from whole blood by Ficoll gradient separation (GE Healthcare, Little Chalfont, UK) and divided into 2–3 aliquots. Sera were separated from whole blood by centrifugation. An aliquot of PBMC and an aliquot of serum were subjected to real-time polymerase chain reaction (PCR) to detect and quantify HHV-6 DNA. Briefly, DNA was isolated from PBMC or serum using the QIAamp DNA blood mini kit (QIAGEN, Hilden, Germany), according to the manufacturer’s protocol. Real-time PCR was performed with the TaqMan fast advanced master mix (Applied Biosystems, Foster City, CA, USA) and the following primers and probe (3): forward primer: GAAGCAGCAATCGCAACACA, probe: AACCCGTGCGCCGCTCCC, reverse primer: ACAACATGTAACTCGGTGTACGGT. The PCR and data collection were conducted on an Applied Biosystems StepOnePlus real-time PCR system. A further aliquot of PBMC was subjected to magnetic bead purification (Miltenyi Biotec, Bergisch Gladbach, Germany) to obtain CD14+ cells. The rest of the cell fraction was subsequently used to purify the CD4 T-cell fraction. The HHV-6 DNA load of each cell type was then measured by real-time PCR. In some experiments, a further aliquot of PBMC was subjected to CD16+ cell isolation followed by CD8 T-cell isolation using magnetic beads. To detect the U31, U39, U90, and U94 gene transcripts, purified CD4 T cells from PBMC were cultured with 5 µg/ml phytohaemagglutinin (PHA) and 20 units/ml recombinant human interleukin 2 in GIT medium (WAKO, Tokyo, Japan). Seven days later, the cells were harvested and subjected to RNA extraction using an RNeasy plus kit (QIAGEN) followed by cDNA synthesis using a high-capacity RNA-to-cDNA kit (Applied Biosystems). Real-time PCR was carried out using specific primers and probes.
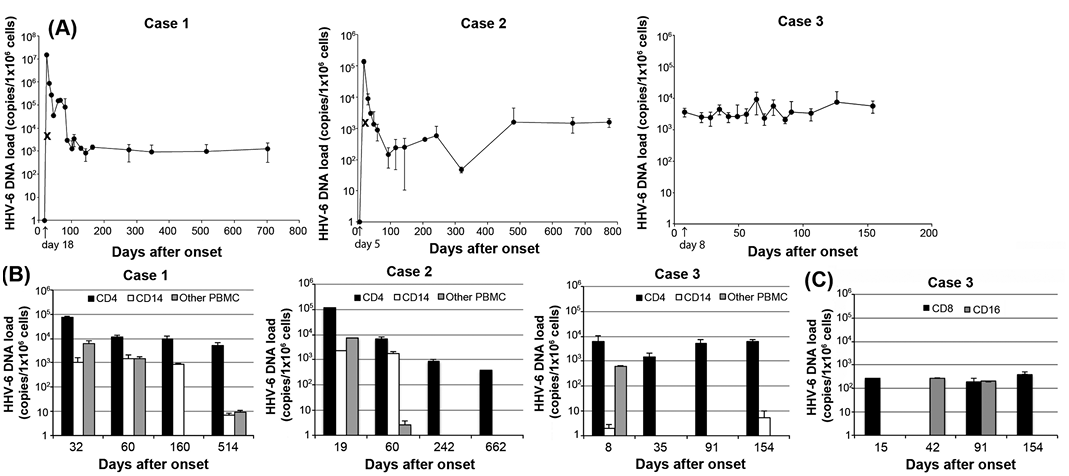
As shown in Fig. 1A, HHV-6 DNA was detected at relatively high copy numbers long after resolution of the patients’ DIHS, although the amounts of DNA detected at these time-points were lower than those seen during the early phase of the condition, except in case 3, in which HHV-6 DNA was detected on the day of admission (day 8).
Fig. 1. Human herpesvirus 6 (HHV-6) DNA load (log scale) in peripheral blood mononuclear cells (PBMC). (A) Real-time PCR was performed with PBMC obtained from 3 drug-induced hypersensitivity syndrome (DIHS) patients. Triplicate experiments, results presented as mean and standard deviation. The arrow indicates the day of admission (when the first sample was collected). × indicates the serum HHV-6 DNA load (copies/ml). HHV-6 DNA was only detected in the patients’ sera when the HHV-6 DNA load of the PBMC was at its peak. (B) Real-time PCR was performed with purified CD4 T cells, CD14+ cells, and the other cells left after the isolation of CD4 T cells and CD14+ cells from the PBMC. The CD4 T cells exhibited a higher HHV-6 DNA load than the CD14+ cells and the remaining PBMC in the early phase. Later, HHV-6 DNA was mainly detected in CD4 T cells. (C) Real-time PCR was performed with purified CD8 T cells and CD16+ cells obtained from case 3.
Since little is known about which types of PBMC harbour HHV-6 in DIHS patients with persistent HHV-6 infections, we next evaluated the HHV-6 DNA loads of CD4 T cells, CD14+ cells, and the remaining PBMC obtained from the 3 patients. During the early phase of the patients’ DIHS, HHV-6 DNA was detected in all cell types, with CD4 T cells being the predominant cell type. At later time-points, CD4 T cells seemed to harbour the majority of the HHV-6 DNA load (Fig. 1B).
HHV-6 was found to mainly infect and replicate in CD4 T cells. However, HHV-6 is able to infect a wide variety of cell types, including natural killer cells and dendritic cells (4). In the latent state, HHV-6 is reported to persist in monocytes/macrophages (4). In some cases, HHV-6 DNA could not be detected in PBMC from healthy individuals with latent HHV-6 infections (4), whereas in others low levels of HHV-6 DNA (around 2 log10 copies/ml) were detected (5, 6).
In our study, no HHV-6 DNA was detected in the patients’ sera at later time-points (Fig. 1A), and while the patients’ anti-HHV-6 IgG titres increased during the early stages of their conditions they subsequently started to decline (Table SI), which is not consistent with reactivation. These findings suggest that latent HHV-6 persisted in the patients’ CD4 T cells at later time-points. However, the amounts of HHV-6 DNA and the types of cells harbouring HHV-6 DNA (CD4 T cells) at later time-points cannot be fully explained by a latent infection. To distinguish between HHV-6 reactivation and latency at later time-points, we examined the expression of 4 HHV-6 gene transcripts, U31, U39, U90, and U94, in CD4 T cells that had been stimulated with PHA. U31 and U39 encode HHV-6 late proteins. U90 is an immediate early gene transcript, and U94 is a putative latency-associated gene transcript (7). Real-time PCR showed that the U90 gene transcript, but not the other 3 gene transcripts, was detected at a low level in CD4 T cells from case 3 on day 77 after onset (data not shown). These results suggest that low-level HHV-6 reactivation that did not have an influence on the viral antibody titre persisted in case 3.
It has been reported that the number of circulating monomyeloid precursors with the CD11b+CD13+CD14–CD16high phenotype increases in the early stages of DIHS and that these cells harbour the HHV-6 antigen (8). Thus, we examined the expression of HHV-6 DNA in CD16+ cells in case 3 (Fig. 1C). Although we were able to detect small amounts of DNA in the CD16+ cells as well as in the CD8 T cells, CD4 T cells served as the main HHV-6 reservoir throughout the course of the patient’s DIHS (Fig. 1B).
Why is HHV-6 DNA persistently detected in some cases of DIHS? HHV-6 infections are frequently encountered in immunosuppressed patients, such as bone marrow transplant recipients (9) and patients with AIDS (5). In DIHS, reductions in immunoglobulin levels and B-cell counts (10), and the marked expansion of functional regulatory T cells (11) have been considered to cause immunosuppression. However, these immunosuppressive conditions occur only in the acute stage of DIHS, which could facilitate HHV-6 reactivation (10, 11). It remains unclear whether our patients were in an immunosuppressed state even after their recovery.
Occasionally, healthy individuals have been shown to have persistently high HHV-6 DNA levels originating from chromosomally integrated HHV-6 (6). The whole-blood HHV-6 DNA levels of immunocompetent individuals that have undergone chromosomal HHV-6 integration are characteristically high (around 6 log10 copies/ml) (6). We investigated cases 2 and 3 for HHV-6 chromosomal integration by examining the patients’ hair follicles. HHV-6 DNA was not detected in their hair follicles (data not shown), indicating that HHV-6 chromosomal integration had not occurred in these cases. Although the possibility of HHV-6 chromosomal integration was not investigated in case 1, the fact that no HHV-6 DNA was detected at the initial visit (day 18) and relatively low levels of HHV-6 DNA were detected at later time-points suggest that case 1 did not involve chromosomally integrated HHV-6.
The authors thank Ayako Yamamoto for her technical assistance.
This study was supported in part by Health and Labor Sciences Research Grants (Research on Intractable Diseases) from the Ministry of Health, Labor, and Welfare of Japan and a Grant-in-Aid for Scientific Research (KAKENHI) to H.A.


 Click to show fullsize
Click to show fullsize