


1Department of Dermatology, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430022, China, 2Department of Dermatology, Affiliated Hospital of Nantong University, Nantong, China, and 3Department of Pharmacology, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, China
#These authors contributed equally to this work.
Epigenetics plays an important role in the development and progression of many diseases. There is increasing evidence for the importance of epigenetic modifications in the progression of psoriasis. The aim of this study was to examine the role and potential mechanism of action of 5-hydroxymethylcytosine (5-hmC) and ten-eleven translocation-2 (TET2) in psoriasiform dermatitis in mice. Immunohistochemical staining was performed on psoriasis patients and healthy controls. Topical application of imiquimod cream to the dorsal skin of mice was used to induce psoriasiform dermatitis. In comparison with healthy controls, 5-hmC was more extensive and intense in the skin lesions from psoriasis patients. TET2 and 5-hmC were highly expressed in imiquimod-induced psoriasiform skin lesions. Importantly, knockdown of TET2 expression in mice attenuated the psoriasiform phenotype and the expression levels of proinflammatory cytokines (interleukin-17A and -17F and interferon-γ) and the chemokine CXCL1 in the lesional skin of mice. This is the first demonstration of a critical role for TET2 in psoriasiform dermatitis in a mouse model, and indicates that 5-hmC may serve as a potential biomarker of psoriasis.
Psoriasis is one of the most common chronic inflammatory skin diseases which is a complex disorder involving multiple pathogenic factors. This study explored the role of epigenetic modifications in the pathogenesis of psoriasis. Our study found that 5-hydroxymethylcytosine and ten-eleven translocation-2 (TET2) levels were increased in psoriatic lesions, compared with normal skin. Knockdown of TET2 expression alleviated the severity of imiquimod-induced psoriasiform dermatitis in mice. Our data highlight a possible role for TET2 in the pathogenesis of psoriasiform dermatitis.
Key words: ten-eleven translocation 2; 5-hydroxymethylcytosine; psoriasis; epigenetics; Th17 cell.
Accepted Mar 15, 2018; Epub ahead of print Mar 15, 2018
Acta Derm Venereol 2018; XX: XX–XX.
Corr: Hongxiang Chen, Department of Dermatology, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Jiefang Avenue 1277#, Wuhan 430022, China. E-mail: hongxiangchen@hotmail.com
Psoriasis is a common chronic inflammatory skin disease associated with well-demarcated, erythematous plaques and silvery scales (1). The pathogenic manifestations of psoriasis are characterized by immune cell infiltration and the aberrant proliferation and differentiation of epidermal cells (2). Although the aetiology of psoriasis is incompletely understood, the interplay between genetic and environmental triggers is known to have an important role in the development of psoriasis (3). Some studies have suggested that abnormal DNA methylation is present in psoriatic skin compared with normal or uninvolved psoriatic skin (4, 5). However, DNA hydroxymethylation and the mechanisms of psoriasis remain poorly understood.
Epigenetic modifications, such as DNA methylation, DNA hydroxymethylation and histone modifications, are crucial for regulating the occurrence and development of a variety of skin diseases, such as systemic lupus erythematosus. This suggests a significant role for epigenetic regulation in dermatology (6, 7). Recently, several studies in both human and mice have identified that the ten-eleven translocation (TET) family dioxygenases TET1, TET2 and TET3 oxidize 5-methylcytosine (5-mC) to 5-hmC, 5-formylcytosine (5-fC) and 5-carboxylcytosine (5-caC) in DNA, and promote DNA demethylation (8–10) . The newly characterized 5-hmC is commonly differentially expressed in various types of disorders. TET2 is expressed in several differentiated tissues and has been studied in a variety of diseases, such as haematopoietic diseases, immune system disorders and cancer (11–14). TET2 has been positively associated with 5-hmC and regulation of other elements (15). Dysregulation of TET2 affects diverse biological processes, including cell differentiation, development, and proliferation, indicating that TET2 is a key epigenetic regulating factor.
Although TET2 functions as a tumour suppressor within the hematopoietic system (13), it is unclear whether TET2 is involved in the pathogenesis of psoriasis. TET2 has been shown to be associated with enriched areas of 5-hmC and to promote signature cytokine expression patterns of Th1 and Th17 cells (12). TET2 selectively mediates transcription of IL-6 during inflammation resolution in dendritic cells by recruiting Hdac2 (16). A previous study showed that some regions of the genome in psoriasis patient T cells displayed prominent hypomethylation compared with the DNA of a healthy control group (17). Whether TET2 and hydroxymethylation could influence immune inflammatory reactions and psoriasis progression is not known.
In the current study, psoriasiform dermatitis was induced in mice using imiquimod (IMQ), which is a classic animal model of psoriasis (18). The impact of knockdown of TET2 expression on the inflammatory response and progression of psoriasiform dermatitis were explored. TET2 was found to maintain the proinflammatory response and induce cell proliferation, promoting progression of psoriasiform dermatitis in the mouse model.
Female BALB/c mice were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd (Bejing, China). The experimental mice were 18–20 g in weight. All experimental protocols with animals were conducted following the ARRIVE guidelines (19). The procedures were approved by the ethics committee for experimental animals, at Tongji Medical College, Wuhan, China.
BALB/c mice were randomized into 3 groups: (i) control group (n = 8), mice were topically treated with Vaseline® and received a subcutaneous injection of negative control shRNAs (lentivirus-delivered scrambled RNAs (sh-sc)) on days 1 and 4; (ii) IMQ group (n = 8), skin inflammation was induced by treatment with IMQ, as reported previously (18). Mice received a daily topical dose of 62.5 mg 5% IMQ cream (Mingxin Pharmaceuticals, Sichuan, China) on shaved backs for 7 consecutive days and a subcutaneous injection of the same dose of sh-sc; (iii) IMQ+sh-TET2 group (n = 8), mice received a subcutaneous injection of lentiviral shRNAs targeting TET2 (sh-TET2, GeneChem, Shanghai, China) followed by the application of IMQ cream for 7 days. On day 8, skin tissue, spleen and blood samples were taken for further experiments.
An objective scoring system was used to score the severity of inflammation on the back skin based on clinical Psoriasis Area and Severity Index (PASI). Erythema, scaling and thickening were classified as follows: 0, none; 1, slight; 2, moderate; 3, marked; and 4, very marked. The sum of these 3 indicator scores served as a measure of the severity of inflammation.
Samples were lysed with cell lysis buffer, and the supernatant was collected by high-speed centrifugation (12,000 rpm). The protein concentration was measured using a bicinchoninic acid (BCA) protein assay kit. Equal quantities of protein were separated by sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE). Protein levels were analysed by specific primary antibodies against TET2 (1:1000; Abcam, Cambridge, USA) or glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (1:1000; Cell Signaling Technology, Massachusetts, USA), which were incubated with the blots at 4°C overnight, followed by incubation with the appropriate horseradish peroxidase (HRP)-conjugated secondary antibodies for 1 h at room temperature. Signals were detected using the DNR bio-imaging systems (Neve Yamin, Israel) according to the manufacturer’s instructions. TET2 protein expression was normalized to that of GAPDH using ImageJ software.
Sections from mouse skin samples, which had been fixed in formalin and embedded in paraffin, were stained with haematoxylin and eosin. Images were captured using a light microscope. The epidermal thickness was measured using ImageJ software. The mean thickness value was calculated by selecting 4 random fields of view in every section.
For immunohistochemical analysis, after routine dewaxing and hydration, the sections were treated with endogenous peroxidase. The pressure cooker antigen retrieval method was used, and the sections were then incubated overnight at 4°C with primary antibodies against 5-hmC (1:1000; Active Motif, California, USA), Ki67 (1:500; Servicebio, Wuhan, China), PCNA (1:1000; Servicebio) and K10 (1:3000; Abcam, USA). The sections were incubated for 30 min with biotin-linked secondary antibodies and then developed with diaminobenzidine. Analysis was carried out using a fluorescent microscope.
Total RNA was extracted from whole skin samples using TRIzol Reagent (Invitrogen, California, USA) according to the manufacturer’s protocol. The RNA concentration was determined by absorbance spectroscopy (NanoDrop, Thermo Scientific Massachusetts, USA), and cDNA was synthesized by a reverse-transcriptase reaction with the PrimeScript RT Reagent Kit (Takara Biotechnology, Ohtsu, Japan). The specific cDNA fragments were amplified using a Real-time PCR Detection System and Power SYBR Green (Takara Biotechnology, Ohtsu, Japan) with the appropriate primers. The data were recorded as expression relative to that of the GAPDH gene. The PCR primer sequences are shown in Table SI.
Genomic DNA from the mouse skin lesions was isolated using the DNeasy Blood & Tissue Kit (Qiagen, Hilden, Germany) according to the manufacturer’s protocol. DNA concentration was then determined by absorbance spectroscopy. The percentage of 5-hmC in total genomic DNA was measured using an ELISA kit (Active Motif) according to the manufacturer’s instructions. All assays were performed in duplicate.
Genomic DNA was purified and sonicated using the above-mentioned DNA extraction kit. The dot blot assay was performed as described previously (20). Briefly, equal amounts of DNA samples were spotted on a nylon membrane. After ultraviolet crosslinking for 10 min, the membrane was blocked with 5% milk in Tris Buffered Saline with Tween for 1 h. The membrane was then incubated with the anti-5-hmC antibody (1:10000; Active Motif) overnight at 4°C. After washing, the membrane was incubated with HRP-conjugated secondary antibodies for 1 h at room temperature. Detection was performed using the DNR bio-imaging systems. The densitometry quantification analysis was completed using ImageJ software.
Individual cell suspensions from the spleen were isolated using a 70-μm cell strainer. The splenocytes were washed twice with PBS and then seeded into 24-well plates, and 1×106 cells/well were cultured with RPMI 1640 medium (HyClone, Utah, USA) with 10% foetal bovine serum. After incubation with Leukocyte Activation Cocktail with BD GolgiPlug (BD Biosciences, New Jersey, USA) for 4 h, the splenocytes were stimulated with Anti-Mouse APC-CD4 in the dark for 30 min at room temperature and then incubated with the Cytofix/Cytoperm Solution Kit (BD Biosciences). Anti-Mouse PE-IL-17A (BD Biosciences) was added to the cell suspension in each tube for 30 min in the dark. Data analysis was performed using a flow cytometer.
Comparisons between 2 groups were performed using two-tailed Student’s t-test. In some analyses, one-way analysis of variance (ANOVA) was used as indicated (3 groups). The data were expressed as the mean ± standard deviation (SD), and p-values < 0.05 were considered statistically significant. GraphPad Prism v.6.0 software (GraphPad Software, San Diego, CA, USA) was used for statistical analysis.
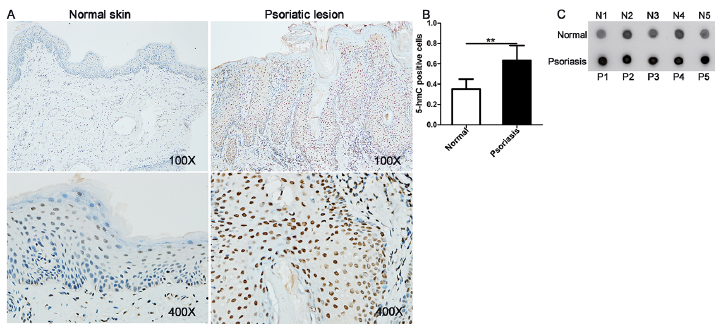
Although aberrant levels of global DNA methylation have been observed in the lesional skin of patients with psoriasis compared with skin from controls (21), the role of hydroxymethylation in the pathogenesis of psoriasis is unknown. Immunohistochemical staining and dot blot assay of 5-hmC were used to determine DNA hydroxy-methylation levels in the skin tissues of patients with psoriasis. In comparison with healthy controls, 5-hmC-positive nuclear staining was more extensive and intense in the skin lesions from patients with psoriasis (Fig. 1A, B), and the expression of 5-hmC was increased in the psoriatic lesions (Fig. 1C). In addition, the mRNA expression levels of TET2, a methylcytosine dioxygenase that catalyses the conversion of 5-mC to 5-hmC, were detected in this earlier study. TET2 expression was found to be upregulated in the psoriasis group (Fig. S1). Therefore, 5-hmC may serve as an epigenetic marker of psoriasis development.
Fig. 1. Abnormal DNA hydroxymethylation in skin lesions of patients with psoriasis. Representative photomicrographs of: (A) psoriatic skin samples and healthy skin samples stained with anti-5-hydroxymethylcytosine antibody. 5-hydroxymethylcytosine (5-hmC)-positive staining was distributed among nuclei throughout the skin. (B) 5hmC-positive nuclei were counted. (C) For quantification, dot blot analysis was conducted. The results are shown as mean±standard deviation (SD). *p < 0.05 and **p < 0.01, compared with the normal samples group.
TET2 is a key methylation-regulating enzyme that can alter DNA methylation status by converting 5-mC to 5-hmC. Therefore, to investigate the possible role of 5-hmC and TET2 in psoriasis pathogenesis, we induced a mouse model of psoriasiform dermatitis by local smearing of IMQ. Simultaneously, shRNAs targeting TET2 or negative control shRNAs (sh-sc) were injected subcutaneously according to the established time point (Fig. 2A). IMQ treatment altered the mouse dorsal skin phenotype after 2 days, and the skin lesions gradually increased in severity over the next 6 consecutive days. The phenotype of IMQ-induced psoriasiform dermatitis was notably ameliorated by administration of sh-TET2, compared with negative control shRNAs (Fig. 2B). The clinical PASI for erythema, scaling, and thickness were markedly decreased in the IMQ+sh-TET2 group compared with the scores in the IMQ group. The differences in scores were statistically significant starting at day 4. Mice in the control group that were treated with Vaseline® and control shRNAs showed no significant changes in their cumulative PASI scores (Fig. 2C).
Fig. 2. Knockdown of TET2 alleviated the morphological and histological changes associated with imiquimod (IMQ)-induced psoriasiform dermatitis. (A) Schematic representation of the animal experiment protocol. (B) Representative images of the dorsal skin of BALB/c mice following continuous treatment at day 8. (C) Epidermal erythema, scaling and thickening of the dorsal skin lesions were evaluated daily. The sum of these 3 criteria scores served as the Psoriasis Area and Severity Index (PASI) score (range 0–12). (D) The epidermal thickness of the dorsal skin was analysed at day 8. (E) Skin biopsies from the dorsal skin of the 3 groups of mice were stained with haematoxylin and eosin (H&E). Scale bar: 100 μm. (F, G) Protein levels of TET2 were analysed by western blotting. (H) mRNA expression of TET2 was analysed by real-time quantitative PCR in skin biopsies from the mice. *p < 0.05, **p < 0.01 and ***p < 0.001, compared with the mice in the IMQ group. Each bar represents the mean ± standard deviation (SD) (n = 6). The data represent the mean of 3 independent experiments, each performed in triplicate.
Histological analyses of the mice from the IMQ group revealed epidermal hyperplasia, acanthosis, parakeratosis, telangiectasia, and abundant inflammatory cell infiltration. Histological changes were strikingly ameliorated in the sh-TET2-treated mice (Fig. 2E). The mean epidermal thickness was measured. Compared with the control mice (24.62 ± 1.10 μm), the epidermal thickness was significantly increased in the IMQ group (206.5 ± 32.38 μm). The epidermal thickness was clearly reduced in sh-TET2-treated mice (52.70 ± 2.296 μm), but still higher than in the control mice (Fig. 2D). Previous studies have indicated that TET2 acts as a key epigenetic regulator (11, 12). We next explored the expression levels of TET2 in the lesional skin of mice. The protein levels of TET2 were examined by western blotting in the skin homogenates, and the results showed that TET2 expression was increased in the IMQ group. However, this expression was significantly suppressed by sh-TET2 treatment (Fig. 2F). The quantitative density value of the TET2 protein was normalized to that of GAPDH, as shown in Fig. 2G. The real-time RT-PCR results were in line with the western blot findings, as the relative mRNA expression of TET2 was markedly upregulated in the IMQ group, compared with that of the control group, and downregulated following injection of shRNAs targeting TET2 into the dorsal skin (Fig. 2H). Thus, the increased expression of TET2 in IMQ-induced psoriasiform dermatitis was visibly reduced by sh-TET2 treatment. These results indicate that TET2 may play an important role in the induction of psoriasiform dermatitis in mice.
TET2 is a dioxygenase that catalyses the conversion of 5-mC into 5-hmC, and diminished TET2 expression or activity is correlated with the expression levels of genomic 5-hmC and subsequently affects the progression and outcome of numerous diseases (10, 12). Therefore, the current study investigated 5-hmC expression in the lesional skin of mice. The distribution of 5-hmC in mouse skin lesions was detected by immunohistochemistry. As shown in Fig. 3A, 5-hmC-positive staining was apparent in nuclei from both the epidermis and dermis cells of the 3 groups. The 5-hmC expression levels in the IMQ group were high, whereas those in the control group and the IMQ+sh-TET2 group were low. Quantification of global 5-hmC levels in the skin lesions was performed, and the results showed that the percentage of 5-hmC was increased in the IMQ-treated mice, but this overexpression was decreased by pretreatment with sh-TET2 (Fig. 3B). In addition, a dot blot assay, which provides a semi-quantitative analysis of 5-hmC levels, was performed, and global 5-hmC levels in genomic DNA from the IMQ-treated mice were increased 2-fold relative to the mice that did not receive IMQ treatment. Consistent with the decrease in TET2 expression, 5-hmC levels were significantly decreased following pretreatment with sh-TET2 (Fig. 3C, D). These data indicate that abundance of 5-hmC in psoriasiform dermatitis is significantly modulated by TET2 expression.
Fig. 3. TET2 as a methylation-regulated enzyme modulated the expression of 5-hydroxymethylcytosine (5-hmC) in imiquimod (IMQ)-induced psoriasiform dermatitis. (A) Immunohistochemical staining for 5-hmC in the dorsal skin samples of controls, IMQ, and IMQ+sh-TET2 mice (n = 6). Scale bar: 100 μm. (B) The percentage of 5-hmC in total genomic DNA were measured using the global 5-hmC quantification assay. (C, D) Dot blot analysis for 5-hmC was conducted in skin biopsies from the mice. The results are shown as mean ± standard deviation (SD). *p < 0.05, **p < 0.01 and ***p < 0.001, compared with the mice in the IMQ group. The data represent the mean of 3 independent experiments, each performed in triplicate.
To understand the TET2-mediated histological alterations that occur in psoriasiform dermatitis, immunohistochemical analysis was performed on skin lesions isolated from mice. The expression of phenotypic markers relevant to proliferation (Ki67 and PCNA) and differentiation (K10) were analysed in each group of samples. Immunostaining for Ki67 in the nucleus revealed a markedly increased proportion of positively stained cells in both the basal and suprabasal epidermal layers following IMQ treatment. However, this staining pattern was obviously suppressed by sh-TET2 treatment (Fig. 4A). Mouse skin samples from both the control group and the sh-TET2-treated group exhibited PCNA-stained nuclei primarily in the basal layer of the epidermis, whereas PCNA-positive nuclei were found in nearly all of the epidermal layers in the IMQ group (Fig. 4B). Immunohistochemical analysis of the differentiation marker K10 revealed an intense staining pattern among all epidermal layers in the control skin, whereas a postponed and lighter staining pattern was observed following IMQ treatment. A moderate increase in K10 staining was observed following sh-TET2 pretreatment (Fig. 4C). These data indicate that keratinocyte proliferation and differentiation are impacted by TET2 expression.
Fig. 4. The effect of TET2 on the expression of Ki67, PCNA and K10 in the epidermis of psoriasiform dermatitis. Representative photomicrographs of the dorsal skin samples of controls, imiquimod (IMQ), and IMQ+sh-TET2 groups stained with (A) anti-Ki67 antibody, (B) anti-PCNA antibody, and (C) anti-K10 antibody. Scale bar: 100 μm.
TET2 and hydroxymethylation may influence immune inflammatory reactions in some diseases. To elucidate the major modulatory effect of TET2 on the pathogenic mechanisms of psoriasis, the expression of RORγt and T-bet, which are key transcription factors involved in Th17 and Th1 cells differentiation, respectively, were investigated. The real-time RT-PCR data indicated a lower expression of RORγt and T-bet following administration of sh-TET2 in the lesional skin of mice, compared with the IMQ group (Fig. 5A, B). Ly6G, a marker of monocytes and granulocytes, correlated with the differentiation and maturation of these cells. As shown in Fig. 5C, Ly6G mRNA expression was upregulated by IMQ treatment and clearly downregulated by sh-TET2 treatment. Ly6G was mainly distributed in the dermis (Fig. S2). In addition, compared with the IMQ group, the increased percentage of Th17 cells was reversed by sh-TET2 treatment in the spleen (Fig. S3). The production of proinflammatory cytokines and chemokines in the skin lesions was investigated. As shown in Fig. 5D–J, the mRNA expression levels of IL-17A, IL-17F, IL-22, IL-23, IFN-γ, CXCL1 and CCL2 were notably increased in the IMQ-treated group compared with those in the control group. However, knockdown of TET2 attenuated the mRNA expression levels of these cytokines and chemokines, which were induced by IMQ treatment, except for IL-22, IL-23 and CCL2. Moreover, some proinflammatory cytokines associated with psoriasis development were detected in the mouse serum samples by cytometric bead array. The increased concentrations of serum IFN-γ, IL-6 and TNF-α were obviously decreased by pretreatment with sh-TET2, but were not completely reverted to control levels (Fig. S4). These findings suggest that TET2 may accelerate IMQ-induced inflammation through mechanisms that involve modulation of immune cell populations.
Fig. 5. The effect of TET2 on immune cell differentiation and the expression of key proinflammatory cytokines and chemokines. Real-time quantitative PCR analysis was performed to determine the mRNA expression levels of: (A) RORγt, (B) T-bet, and (C) Ly6G in skin biopsies from mice. (D–H) mRNA expression levels of proinflammatory cytokines (interleukin (IL)-17A, IL-17F, IL-22, IL-23, interferon (IFN)-γ) in skin biopsies from mice. (I, J) Expression of chemokines (CXCL1, and CCL2) mRNA in the back skin. The results are shown as the mean ± standard deviation (SD). *p < 0.05, **p < 0.01 and ***p < 0.001, compared with the mice in the imiquimod (IMQ) group. The data represent the mean of 3 independent experiments, each performed in triplicate.
DNA hydroxymethylation is one of the most pivotal epigenetic regulators of gene expression (22, 23). The TET2 protein is a critical regulator of DNA hydroxy-methylation. In recent years, TET2-induced DNA hydroxymethylation has been associated with critical regulation of the molecular pathogenesis of various diseases (11, 24, 25). Immune responses related to T-helper-cell differentiation are regulated by epigenetic changes (26). Our study found that 5-hmC levels were significantly increased in psoriatic lesions, compared with normal skin, by immunohistochemical analysis. Subsequent studies have indicated that reduction of TET2 expression by knockdown alleviates the severity of IMQ-induced psoriasiform dermatitis in mice. Our data highlight a possible role for TET2 in the pathogenesis of psoriasiform dermatitis.
Th17 and Th1 cells are pivotal effectors of autoimmune inflammation. IMQ-induced psoriasiform skin inflammation is dependent on the presence of T cells (18). IL-17 is an important proinflammatory cytokine produced by Th17 cells. IL-17 can activate various immune cells to produce a variety of cytokines and chemokines, and stimulates the production of antimicrobial peptides that promote recruitment of inflammatory cells (27). The pathogenic mechanisms of psoriasis are critically dependent on the IL-23/IL-17 axis (28). The lineage-specific transcription factors T-bet and RORγt are key regulators of Th1 and Th17 cell function, respectively. In a recent study, TET2 regulated Th1 and Th17 cell differentiation in a T-bet-dependent and RORγt-dependent manner, respectively. TET2 induces DNA hydroxymethylation at the IFN-γ and IL-17 loci and thereby upregulates the expression of inflammatory cytokines in a particular Th cell subset (12). Consistent with these reports, our study showed that knockdown of TET2 expression evidently reduced Th17 cell differentiation and inflammatory factor production (IL-17A, IL-17F, and IFN-γ) in IMQ-induced psoriasiform dermatitis in mice.
Growing evidence has shown that proinflammatory mediators (IL-22, IL-23, IFN-γ, IL-6 and TNF-α) and chemokines (CXCL1 and CCL2) play critical roles in the initiation and aggravation of inflammatory responses in psoriasis (1, 29). Using an integrative bioinformatics approach, CXCL1 was identified as a hub gene in psoriasis because it was aberrantly methylated and expressed compared with normal skin (data not shown). Our results also show that expression of these proinflammatory cytokines and chemokines is increased in the skin lesions or serum of IMQ-treated mice compared with controls. Moreover, the decreased TET2 expression following injection of lentiviruses expressing shRNAs targeting TET2 significantly downregulated the expression of IFN-γ, IL-6, TNF-α and CXCL1, but the expression of IL-23 and IL-22 were not affected. These findings indicate that TET2 may partly enhance some cytokine expression to participate in the progression of IMQ-induced psoriasiform skin inflammation.
The commonly considered pathogenesis of psoriasis is a that of variety of inducible cytokines that lead to activation of dendritic cells and secretion of mediators, such as IL-23 and IL-12, leading to the differentiation of Th17 and Th1 cells. T cells secrete inflammatory mediators that induce keratinocytes hyperproliferation and abnormal expression of chemokines, antimicrobial peptides and cytokines. These factors, in turn, further expand the inflammatory response (1). In the present study, we observed that 5-hmC expression levels were significantly decreased in the skin lesions by pretreatment with sh-TET2, compared with in the IMQ group. The expression of IL-17 and IFN-γ were deregulated, but IL-23 expression was not affected. Based on a recent study, in which TET2 regulated DNA hydroxymethylation at the IFN-γ and IL-17 loci (12), it is plausible to speculate that TET2 mainly targets Th17 and Th1 cells. In addition, DNA hydroxymethylation of keratinocytes may also play a role in the development of psoriasiform dermatitis. Keratinocytes are the main secretory cells of CXCL1 in psoriasis skin lesions (30). TET2 may partially affect the pathogenesis of psoriasiform skin inflammation by regulating the methylation status of CXCL1 in keratinocytes.
Within the past decade, much progress has been made in elucidating the pathological mechanisms of psoriasis; therefore, more novel therapeutic strategies have been applied during clinical treatment. The mechanism and treatment breakthroughs have been accompanied by increasing research on epigenetic modifications in diseases (31). Several studies have found abnormal DNA methylation in psoriatic lesions compared with normal controls. These findings suggest that DNA methylation is involved in the pathogenesis of psoriasis (4, 5). However, the DNA hydroxymethylation associated with psoriasis remains poorly characterized. Our data indicate that the TET2 protein influences IMQ-induced psoriasiform dermatitis through mechanisms that involve DNA hydroxymethylation abundance and dysregulation of the inflammatory reaction, which then affects epidermal keratinocyte proliferation and differentiation. The current study is only a preliminary investigation of this mechanism. These results highlight the need for further research focussing on the specific roles of TET proteins in particular signalling networks. It is necessary to characterize whole-genome DNA hydroxymethylation patterns in psoriatic lesions.
In conclusion, this study investigated the potential role and mechanisms by which TET2 affects the patho-logical mechanism of psoriasiform dermatitis. The results suggest the existence of a DNA hydroxymethylation-mediated regulation of T-cell differentiation and cytokine gene expression in IMQ-induced psoriasiform skin inflammation in mice. TET2 and 5-hmC may play important roles in the pathogenesis of psoriasis.
This work was supported by the National Natural Science Foundation of China (numbers 81673057 and 81502730).
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize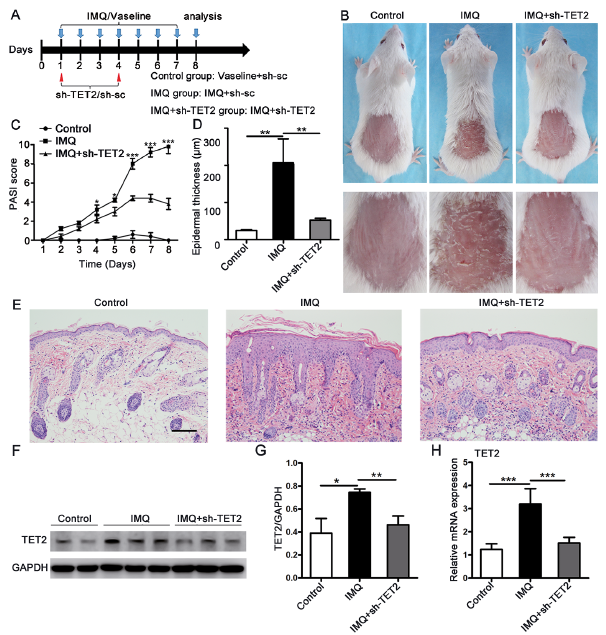 Click to show fullsize
Click to show fullsize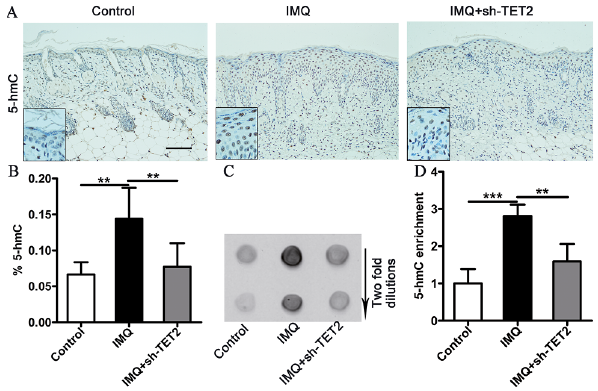 Click to show fullsize
Click to show fullsize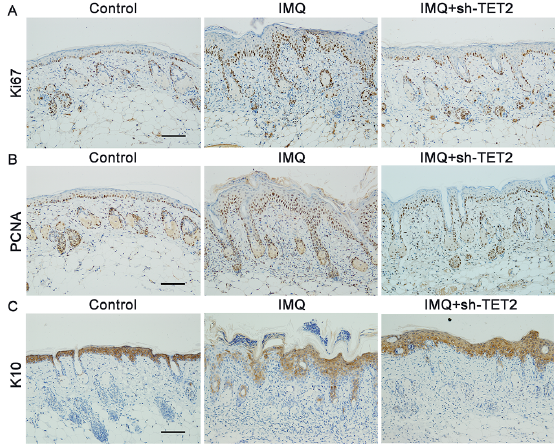 Click to show fullsize
Click to show fullsize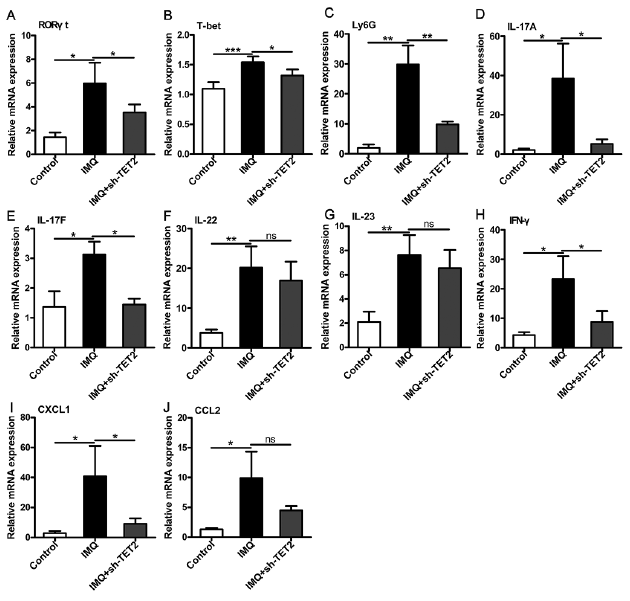 Click to show fullsize
Click to show fullsize