


1Department of Dermatology, Toho University Ohashi Medical Center, Tokyo, and 2Shiseido Global Innovation Center, Yokohama, Japan
#These authors contributed equally and should be considered as first author.
Cells that constitute the dermal papillae of hair follicles might be derived from the dermal sheath, the peribulbar component of which is the dermal sheath cup. The dermal sheath cup is thought to include the progenitor cells of the dermal papillae and possesses hair inductive potential; however, it has not yet been well characterized. This study investigated the gene expression profile of the intact dermal sheath cup, and identified dermal sheath cup signature genes, including extracellular matrix components and bone morphogenetic protein-binding molecules, as well as transforming frowth factor beta 1 as an upstream regulator. Among these, gremilin-2, a member of the bone morphogenetic protein antagonists, was found by in situ hybridization to be highly specific to the dermal sheath cup, implying that gremlin-2 is a key molecule contributing to maintenance of the properties of the dermal sheath cup.
Key words: alopecia; dermal papilla; GREM2; hair biology; TGF-b1.
Accepted Apr 24, 2018; Epub ahead of print Apr 24, 2018
Acta Derm Venereol 2018; XX: XX–XX.
Corr: Shiro Niiyama, Department of Dermatology, Toho University Ohashi Medical Center, 2-17-6 Ohashi, Meguro-ku, Tokyo, 153-8515, Japan. E-mail: sniiyama@aol.com
We investigated the gene expression profile of some part of the hair follicle. To identify the cell signature genes of the hair follicle should contribute the quality control of culturing these cells and could lead to enhance the safety and efficacy on cell-based therapy for hair-related diseases such as androgenic alopecia for both men and women.
Hair follicles (HF) undergo cycles of hair growth, regression and removal. HF are composed of dermis- and epithelium-derived cell populations in close association with each other. The dermal mesenchyme-derived component consists of fibroblast-like cells that form the dermal papilla (DP), located at the centre of the hair bulb, and the dermal sheath (DS), which is the outermost layer of the HF and surrounds the epithelial component (1–4). The DP is regarded as essential to HF development and cycling. Biochemical signalling by DP cells controls the cell dynamics of the epithelial component and the overall physical properties of the HF. It has been shown that isolated DP or even cultured DP cells can induce the formation of new HF following transplantation into the skin (5–11).
It has been suggested that the cells that constitute the DP may be derived from the DS (12). When the bulb regions of rat vibrissae, including the DP, were removed by microdissection, the amputated follicles were shown to regenerate a new DP and renew production of a hair fibre. However, when more than the lower third of a vibrissa follicle was removed, the amputated vibrissa failed to form a new DP or produce a hair fibre (13). By contrast, a new DP was formed when the lower half of a vibrissa follicle was removed and DS tissue isolated from the base of another follicle was implanted into the amputated end (14). These results suggest that the DS may contain DP progenitor cells. Recently, in vivo lineage tracing in transgenic mice has been used to show that the DS contains a pool of mesenchymal stem cells that can replenish DP and DS cell populations lost during the catagen and telogen stages of the hair cycle (15).
McElwee et al. (16) demonstrated that murine DP-derived cells and cells from the peribulbar DS ‘’cup’’ (DSC) lateral to the DP induce new HF in both implanted ears and footpads. DSC cells are distinct from non-bulbar upper DS (UDS) cells. While a number of studies on the DP at both the biological and molecular levels have been published, describing properties and physiological characteristics, gene expression profiles and 3-dimensional culture characteristics of the DS/DSC, the specific gene expression patterns, have been poorly characterized (17–20). Since it is well known that primary cells in culture change or lose some of their physiological properties during culture passages (21), intact DSC profiling would be essential for a precise characterization of the DSC. This report microdissected 3 different mesenchymal subunits of human HF, the DSC, DP and UDS, and performed microarray analyses of RNA from the intact, isolated tissue to identify DSC signature genes and their regulators.
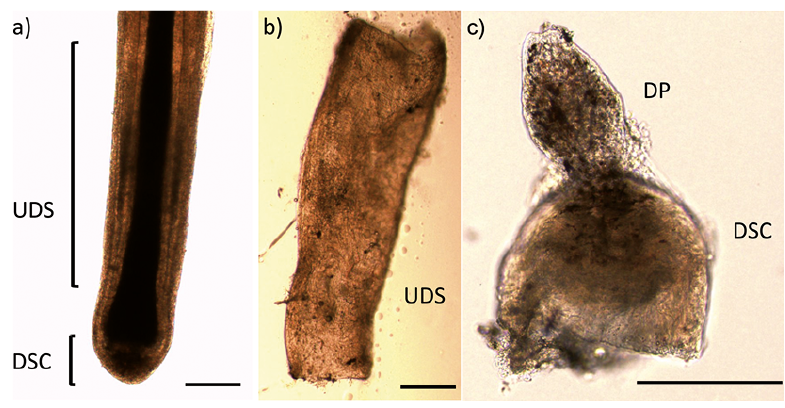
Written informed consent was obtained from all patients participating in this study, which was conducted with the approval of the ethics committee of Toho University Ohashi Medical Center and Shiseido Life Science Research Center. Skin pieces containing scalp HF were obtained from 3 female patients, aged 30–70 years, undergoing excision of 2 cases of melanocytic naevus and one case of lipoma. Tissues were collected from the margin of the excised skin and did not contain nests of these cutaneous tumours. The mesenchymal subunits of HF were microdissected from freshly isolated, intact HF under a stereomicroscope, as described previously (Fig. 1) (16, 22). Briefly, the HF was gripped with forceps immediately adjacent to the bulb region and the bulb was dissected free with a transverse cut using a needle. The DSC of the bulb was inverted using forceps and needles, and any remaining epithelium-derived tissue was removed to expose the DP. The DP was separated from the DSC with a transverse cut. The UDS was isolated by stripping it from the outer root sheath with forceps, using gentle pressure, while the proximal end of the HF was held with another pair of forceps.
Fig. 1. Isolation of the hair follicle (HF) subunits. (a) A whole HF. (b) Dissected upper dermal sheath (UDS). (c) Dissected and inverted dermal sheath cup (DSC) and dermal papilla (DP) before separation. Scale bars: 200 µm.
Microdissected HF tissue from a single subunit and patient was pooled and dissolved in 1 ml ISOGEN (Nippon Gene Co. Ltd, Toyama, Japan), followed by RNA extraction according to the manufacturer’s instructions. After precipitation, RNA quality was assessed from the ribosomal RNA ratio (28S/18S) using an Agilent Bioanalyzer 2100 (Agilent Technologies, Palo Alto, CA, USA). Samples with RNA integrity numbers of 6.5–8.3 were used for microarray analysis.
Microarray analysis was performed according to the manufacturer’s instructions (Agilent Technologies), as described previously (23). Ten nanograms of total RNA was converted to fluorescently labelled cRNA by in vivo reverse transcription, followed by transcription using the Low Input Quick Amp Labeling Kit (Agilent Technologies) in the presence of oligo(dT) primer, AffinityScript reverse transcriptase, T7 RNA Polymerase, and CTP labelled with Cy3 dye. The resulting labelled cRNA (600 ng) was fragmented and hybridized to an Agilent SurePrint G3 Human GE 8×60K Microarray (G4851A, Agilent Technologies), and scanned with an Agilent dual laser microarray scanner (G2565AA). The signal intensities were quantified using Feature Extraction Software Version 11.0.1.1 (Agilent Technologies). For the statistical analysis, the data were processed with Genespring GX 13.0 software (Agilent Technologies). The microarray data were deposited in the Gene Expression Omnibus (GEO) database (accession number GSE95219; http://www.ncbi.nlm.nih.gov/geo) and described in accordance with the Minimum Information About a Microarray Experiment (MIAME) guidelines.
For visualization of the GREM2 localization within cells, in situ hybridization was performed with paraffin-embedded tissue section. A GREM2-specific type 6 probe, based on highly specific, branched DNA signal amplification technology was used according to the manufacturer’s manual for the View RNATM ISH Tissue 1-Plex Assay kit (Thermo Fisher Scientific Inc., Waltham, MA, USA). Signals were detected with Fast-Red substrate, followed by fluorescence view. Nuclear staining was performed by Hoechst 33342 (Thermo Fischer Scientific Inc.).
To determine the physiological characteristics of the hair mesenchymal component in situ in the tissue, freshly isolated, intact HF subunits were analysed (Fig. 1). Microarray analyses of total RNA from the microdissected HF subunits of 3 individual donors were performed to identify HF-subunit-specific signature genes. Microarray probes for which data were tagged as unreliable or equal to background for more than one individual were excluded, leaving 25,342 probes to be analysed. In order to identify intact HF-subunit-specific signature genes, the 18,905 probes for which signal intensity was reproducibly greater than 50 for at least one HF component were selected for further analysis.
Firstly, principal component analysis (PCA) was performed to show the relationship between total microarray datasets. While the locations of the 3 HF subunits from a single individual were associated closely along the PC1 dimension, the locations of the same subunit from different individuals were associated more weakly along the PC3 dimension (Fig. 2). Considering these inter-individual differences, a 1.5-fold cut-off was applied for DSC vs. DP, DSC vs. UDS, and UDS vs. DP comparisons within each individual. PCA showed similarity not only of DSC, but also of DP and UDS profiling among 3 individuals, although there were also substantial inter-individual differences.
Fig. 2. The similarity of microarray results following principal component analysis. Three-dimensional views from different viewpoints. x-axis: principal component (PC) 1 (32.34% variance captured); y-axis: PC2 (22.83% variance captured); z-axis: PC3 (20.43% variance captured). Red symbols: individual (ID) 1; blue: ID2; brown: ID3. Triangles: dermal sheath cup; squares: dermal papilla; circles: upper dermal sheath.
The numbers of up- or down-regulated genes common to all 3 individuals are shown in Table I. A total of 63 genes that were up-regulated in the DSC vs. DP were identified. Among these, the genes that were also up-regulated in the UDS were eliminated. The resultant 32 genes after eliminating the 5 uncharacterized probes, are listed as DSC signature gene list, as shown in Table SI. Similarly to the DSC signature list, 116 DP signature genes are also listed in Table SII.
Table I. Numbers of genes showing expression changes greater than 1.5-fold for each individual considered separately, and for all 3 individuals, in comparisons of the dermal sheath cup (DSC) vs. dermal papilla (DP), DSC vs. upper dermal sheath (UDS) and UDS vs. DP
This DSC signature gene list includes actin, alpha 2, smooth muscle (alpha-smooth muscle actin), prostaglandin E receptor 3 and matrix metallopeptidase 11, which have previously been reported to be expressed in the DS (24–26). Seven extracellular matrix components, decorin, fibulin 2, coiled-coil domain containing 80, tenascin XB, asporin and hyaluronan and proteoglycan link protein 1 and matrix metallopeptidase 11, are also included in the list, and this category was shown to be significantly over-represented in gene ontology analysis (Table II). Most of the DSC signature genes were highly expressed, not only in the DSC, but also in the UDS, compared with the DP (Fig. 3). Among them, 6 genes, asporin, DAN family BMP antagonist GREM2, matrix metallopeptidase 11, prostaglandin E receptor 3, retinol binding protein 4 and transducin-like enhancer of split 4 (TLE4) appear to be more specific to the DSC, showing at least 1.5-fold up-regulation vs. the UDS (Fig. 4, Table SIII).
Table II. Gene ontology (G) analysis of dermal sheath cup (DSC) signature
Fig. 3. Genes differentially expressed in the dermal sheath cup (DSC). Normalized expression levels of DSC signature genes compared with dermal papilla (DP) and upper dermal sheath (UDS) in 3 individuals.
Fig. 4. Genes expression specific to dermal sheath cup (DSC) in dermal papilla (DP) and upper dermal sheath (UDS). Colour legend: red: genes with greater expression relative to the geometric means; green: genes with lower expression relative to the geometric means.
Among 6 DSC specific genes, GREM2 showed substantial up-regulation in DSC, both in comparison with DP and UDS. Therefore, q-PCR for GREM2 was performed with intact DP and DSC tissues and the result clearly showed DSC-specific up-regulation compared with DP, which is almost 10 times higher in DSC than in DP (Fig. 5a). Also, restricted gene expression of GREM2 was localized within the DSC area in the bulb region of HF by in situ hybridization technique, indicating that GREM2 expression is specific to the DSC in intact HF (Fig. 5b–d).
Fig. 5. Expression of GREM2 in human hair follicles. (a) Gene expression level of GREM2 in dermal sheath cup (DSC) and dermal papilla (DP) by q-PCR. In situ hybridization of GREM2: (b) GREM2 probe. (c) High-power field of (b). (d) No probe. Red: in situ signal; blue: nuclear staining by Hoechst 33342. Scale bars: (b) 100 µm, (c, d) 50 µm.
Following analyses of upstream regulators, which evaluate expression changes are consistent with the reported literature, TGF-b1 was identified as a putative regulator of 9 genes of the DSC signature (Table III). Five genes were related to the Wnt pathway, one of which, TLE4, is a member of the Groucho family, which are well-known nuclear components of the Wnt pathway. Two BMP signalling antagonists, GREM1 and GREM2, were also up-regulated in the DSC. Fig. 6 shows the putative regulatory network and subcellular location of DSC signature genes, suggesting that TGF-b1 regulated extracellular matrix components characteristic to DSC.
Table III. Upstream regulators of dermal sheath cup (DSC) signature genes
Fig. 6. Putative regulation and subcellular location of dermal sheath cup (DSC) signature genes. The relation map of DSC upregulated genes was described by Pathdesigner of Ingenuity software (Qiagen, Venlo, Netherlands).
This study compared the gene expression profiles of 3 different mesenchymal subunits of the human HF, the DSC, DP and UDS, which were isolated by microdissection. A DSC signature containing 32 genes was identified, which may be regulated by TGF-b1, Wnt and BMP signalling molecules. Among the DSC signature genes, alpha-SMA, matrix metallopeptidase 11 and prostaglandin E receptor 3 have been reported to be expressed in the DS (24, 26, 27, 28).
Furthermore, this study identified 113 DP signature genes from 127 probes that were up-regulated in the DP compared with the DSC (Table I, Table SII). DP signatures reported previously by 3 groups, and a dermal condensation signature (17–20), respectively, included 18, 12, 19 and 10 genes in common with our results (Table IV). These similarities suggest that the novel signature genes identified are specific to, and characteristic of, each mesenchymal subunit of the HF.
Table IV. Dermal papilla (DP) signature genes common to this study and previous reports
It is noteworthy that TGF-b1 was identified as a putative upstream regulator of the DSC signature genes. TGF-b1 is a well-known regulator of extracellular matrix expression, has previously been reported to be expressed in the DS of human HF, and can regulate the reactivation of DP cells by androgens (29–31). On the other hand, the results of the current study suggest that characteristic extracellular matrix components are expressed in the DSC. In addition, DS cells have been reported to show a distinct profile of matrix metalloproteinases (32, 33). Thus, TGF-b1 may play a role in regulating DSC matrix characteristics. Al-though the origin of the DSC remains unclear, considering that GREM1 and GREM2 are expressed in both mouse embryonic skin and human HF, it is possible that they could play a role in the early stage of DSC formation. In the present study, 3 genes, GREM2, retinoic acid binding protein 4, together with the Wnt signalling molecule TLE4, showed relative higher expression levels in the DSC compared with the DP or DS (Fig. 4, Table SIII). Interestingly, a recent study reported that mesenchymal-derived GREM1 plays a crucial role in feather formation accompanying signalling modulation by Wnt-3a and retinoic acid (34), thus these 3 molecules, including GREM, may be involved in the functioning of the DSC during the hair cycle and hair morphogenesis.
This report identified several signature genes of the DSC and showed specific localization within DSC of one of them, revealing the molecular mechanisms that contribute to acquisition of the characteristic features of the human DSC, which could be useful for understanding the biology of the human HF.
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize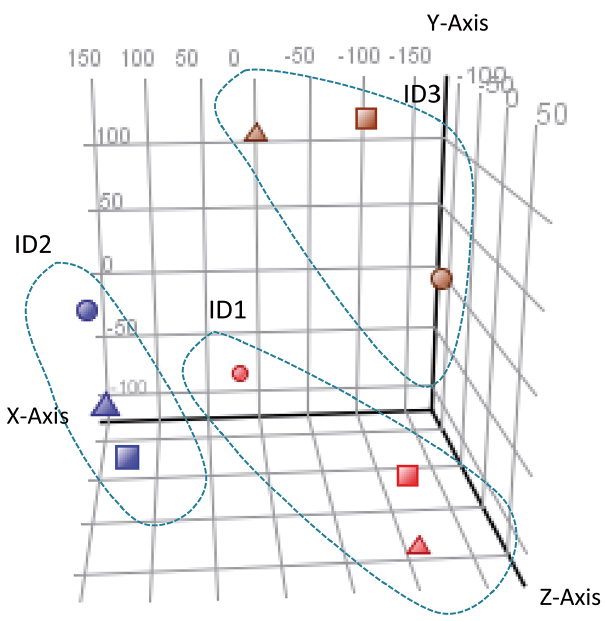 Click to show fullsize
Click to show fullsize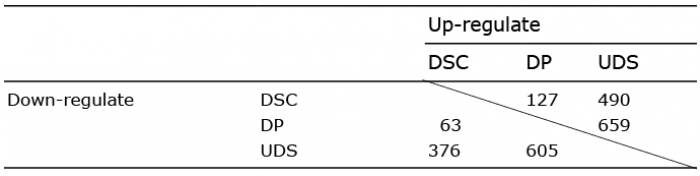 Click to show fullsize
Click to show fullsize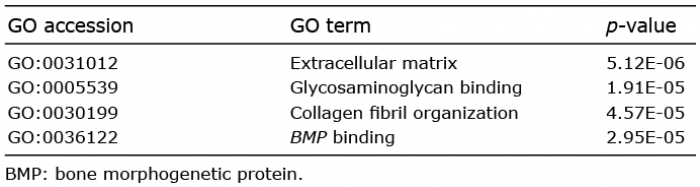 Click to show fullsize
Click to show fullsize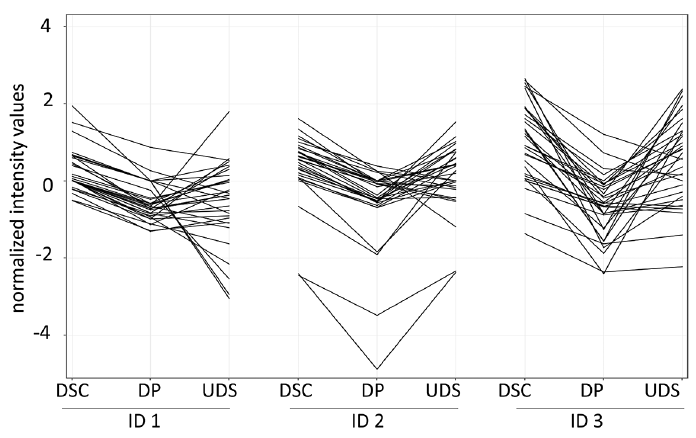 Click to show fullsize
Click to show fullsize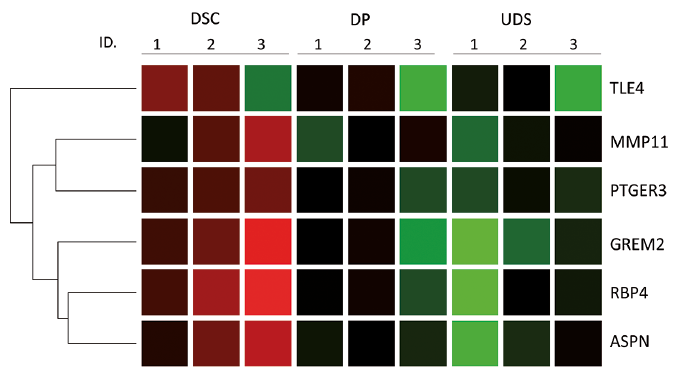 Click to show fullsize
Click to show fullsize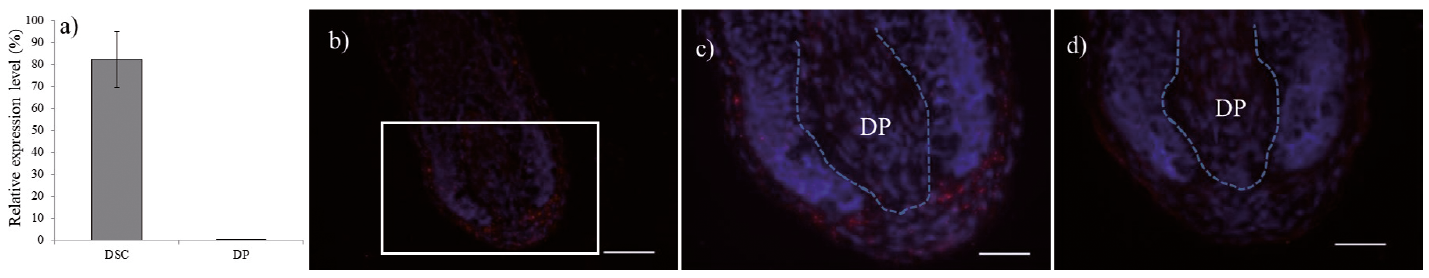 Click to show fullsize
Click to show fullsize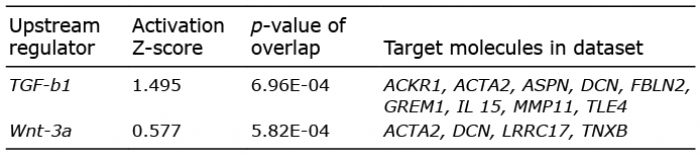 Click to show fullsize
Click to show fullsize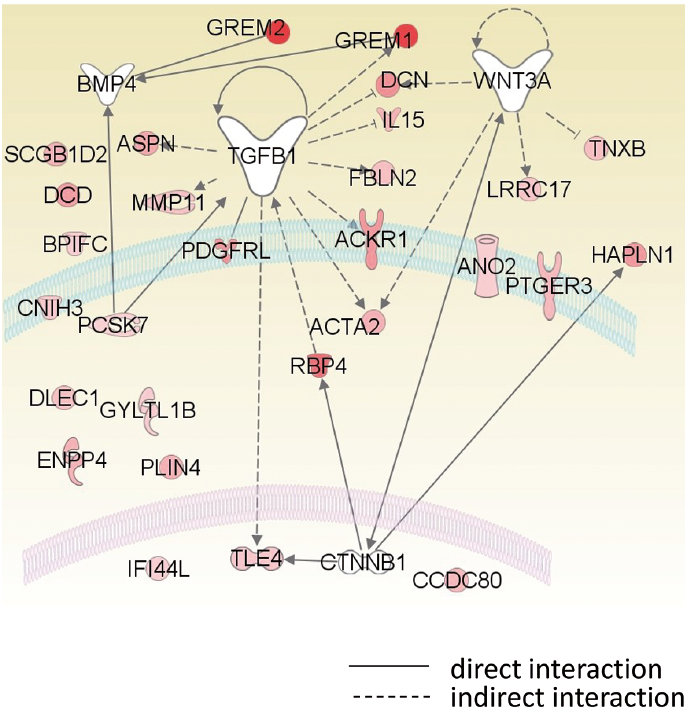 Click to show fullsize
Click to show fullsize Click to show fullsize
Click to show fullsize