


1Lee Kong Chian School of Medicine, 3School of Biological Sciences, 4School of Materials Science and Engineering, 5Nanyang Environment and Water Research Institute, (Environmental Chemistry and Materials Centre), Interdisciplinary Graduate School, Nanyang Technological University, Singapore, 2Epithelial Biology, Institute of Medical Biology, Immunos, 6Skin Research Institute of Singapore, Singapore, 7Wound Healing Center, Bispebjerg University Hospital, Copenhagen, Denmark, and 8Division of Dermatology and Venereology, Department of Clinical Sciences, Lund University, Lund, Sweden. #E-mail: ssadav@ntu.edu.sg
Accepted Apr 24, 2018; Epub ahead of print Apr 24, 2018
Antimicrobial peptides (AMPs) are evolutionarily old components of innate immunity. AMPs identified in human skin include defensins, cathelicidin, dermcidin, psoriasin, and RNase7 (1, 2). Some AMPs, such as the human cathelicidin peptide LL-37, are upregulated during inflammation in skin (1). Various microbes colonize the hair follicular canal, and various AMPs may modulate their population and composition (3, 4). It is not known whether the hair shaft per se contains functionally active antimicrobials.
In a recent study, we explored different extraction procedures to determine the complete proteome of human hair shafts. That work focused on establishing potential biomarkers for hair with importance for evaluating factors, such as ageing, infection, and potentially underlying follicular disorders. A multitude of potential antimicrobial peptides and proteins were observed, including lysozyme, various S100 proteins, and histones (5). This report focuses on the identification of hair-derived molecules with potential antimicrobial activity. Hair shafts were subjected to an acid-based extraction method (Appendix S1). The extracted material was fractionated using reverse-phase high-pressure liquid chromatography (RP-HPLC), and each fraction was analysed for antimicrobial activity by radial diffusion assay. Potential AMPs were identified by Western blotting combined with high-throughput mass spectrometry coupled with liquid chromatography.
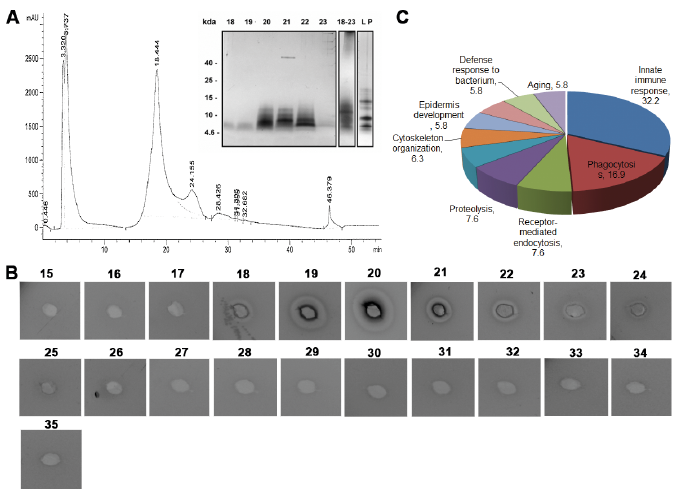
Hair shaft extraction using a buffer containing 8 M. urea (see Appendix S1), followed by mass spectrometric analysis of this extract revealed the presence of multiple proteins (Table SI). Next, we attempted to detect possible antibacterial activity derived from hair shafts. Using RP-HPLC, a major protein peak was observed (Fig. 1A, peak at 18.44, fractions 17–23). The eluent was characterized using SDS-PAGE analysis (Fig. 1A insert) and antibacterial activity was analysed using a radial diffusion assay (RDA) by collecting and analysing fractions eluted between 17 and 23 min (corresponding to 23–40% acetonitrile) (Fig. 1B). The results indicated that the antibacterial activity was related to proteins with molecular masses of 4–15 kDa.
Fig. 1. Profiling and characterization of the low-molecular-weight (LMW) fraction of the human hair proteome. (a) Reverse-phase high-pressure liquid chromatography (RP-HPLC) elution profile of the LMW hair proteome (inset: sodium dodecyl sulphate-polyacrylamide gel electrophoresis of LMW protein extract and individual fractions eluted from minutes 15 to 35. (b) Antimicrobial activity (visualized as bacterial inhibition zones around the 3-mm wells) of individual HPLC fractions as determined by radial diffusion assay (RDA) using Escherichia coli. (c) Gene ontology analysis of the proteins identified in fractions 18–23 (employing PANTHER, Appendix S1).
The RDA analysis demonstrated that fractions 19 and 20 showed zones of inhibition against Escherichia coli, indicating the presence of antimicrobials (Fig. 1B). The fractions showing antimicrobial activity were selected for further analysis by mass spectrometry. The fractions were found to contain multiple protein sequences (Table SII), in particular those linked to host defence, including multiple histone sequences. The gene ontology analysis is shown in Fig. 1C. One example of an identified histone sequence, H2A, is shown in Fig. 2A.
Fig. 2. Identification of histones in fractions separated by high-pressure liquid chromatography (HPLC). (a) Spectrum of histone H2A identified in the combined HPLC fractions 18–23. (b) Western blot analysis of histone isoforms in individual HPLC fractions displaying antimicrobial activity. Antibodies against H1, H2A, H2B, and H3 were used. Bovine serum albumin (BSA) was used as a negative control, and a low-molecular-weight protein (LMW P) extract from human hair shafts was used as a positive control. (c) Immunohistochemical detection of histone H3 in human hair shafts of a formalin-fixed paraffin-embedded skin section using a rabbit polyclonal anti-histone H3 (tri-methyl K9) antibody. (d) Negative control.
To confirm the mass spectrometry results, RP-HPLC fractions 18–23 were further analysed by Western blot using specific rabbit polyclonal antibodies against histones. The results are presented in Fig. 2B and demonstrate proteins of 24, 17 and 14 kDa, which correspond to histones H1, H2B and H3 and H2A, respectively. To illustrate the presence of one such histone variant, immunohistochemistry analysis using an antibody against H3 was performed. The results indicated that this histone is localized in the cortex of hair shafts (Fig. 2C).
Antimicrobial activity of histones was first reported in 1958 by Hirsch (6), who demonstrated that arginine-rich preparations (later denoted as histones) isolated from calf thymus exerted potent bactericidal activity. Subsequent reports showed that histones H1, H2A and H2B isolated from different species can act as potent antimicrobial agents (7–9). Recombinant human histone H1.2 shows activity against both Gram-positive and Gram-negative bacteria, including drug-resistant strains, such as multidrug-resistant Pseudomonas aeruginosa and methicillin-resistant Staphylococcus aureus (MRSA) (10). Histone H2B is found in human placenta and colonic epithelial cells, but also in the skin of tree frogs (11). Chicken H2B (12) displays antimicrobial activity against both Gram-positive and Gram-negative bacteria. Histones H4 and H3 obtained from calf thymus are active against E. coli, and histone H4 also exerts antimicrobial activity (13). Recombinant histone H4 from human sebocytes inhibited the growth of S. aureus and Propionibacterium acnes. Antimicrobial histones are also major components of neutrophil extracellular traps (14). Hence, given the identification of multiple histones in the antibacterial fractions of the hair extracts makes it plausible that these well-known antimicrobials contribute to the observed activity. However, since other components were also identified, it cannot be excluded that the observed bacterial inhibition is due to contributions from other AMPs, or even non-peptide molecules, such as lipids, possibly co-migrating in the RP-HPLC analysis. The latter possibility is unlikely, as possible antibacterial sebum components should be washed away during the initial washes of the hair shafts with 70% ethanol (see Appendix S1).
In conclusion, the present study demonstrates, for the first time, methods to detect and purify antimicrobial factors, such as histones, from hair shafts. Further studies are warranted to explore whether histones or other AMPs of hair shafts confer a resistance to infection on hair in vivo. Furthermore, due to the cationic nature of histones, the results should encourage further research into whether histones, like many cationic polymeric hair conditioners (15), can contribute to a potential cationic charge density at the hair shaft surface. Finally, our study also shows that human hair shafts could be a new source for the purification of functionally active histones.
This work was supported by the Lee Kong Chian School of Medicine, Nanyang Technological University Start-Up Grant, and the Singapore Ministry of Education [MOE2014-T2-2-043 (Tier 2: ARC9/15) to SKS and MOE2015-T1-001-082 (Tier 1 grant) to AS].


 Click to show fullsize
Click to show fullsize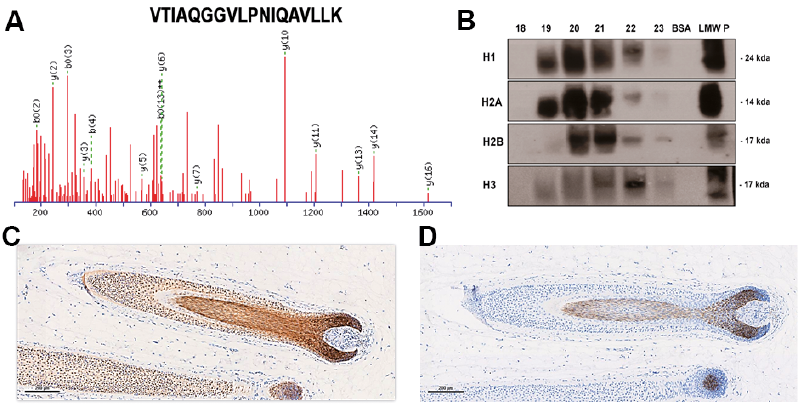 Click to show fullsize
Click to show fullsize