Antioxidative Phytochemicals Accelerate Epidermal Terminal Differentiation via the AHR-OVOL1 Pathway: Implications for Atopic Dermatitis
Masutaka Furue1–3, Akiko Hashimoto-Hachiya1 and Gaku Tsuji1
1Department of Dermatology, 2Research and Clinical Center for Yusho and Dioxin, and 3Division of Skin Surface Sensing, Department of Dermatology, Kyushu University, Fukuoka, Japan
Aryl hydrocarbon receptor (AHR) is a chemical sensor that is expressed abundantly in epidermal keratinocytes. Oxidative AHR ligands induce the production of reactive oxygen species. However, antioxidant AHR ligands inhibit reactive oxygen species generation via activation of nuclear factor-erythroid 2-related factor-2, which is a master switch for antioxidative signalling. In addition, AHR signalling accelerates epidermal terminal differentiation, but excessive acceleration by oxidative ligands, such as dioxins, may induce chloracne and inflammation. However, antioxidative phytochemical ligands induce the beneficial acceleration of epidermal differentiation that repairs skin barrier disruption. The upregulated expression of differentiation molecules, such as filaggrin, is mediated via the AHR-OVOL1 axis. This AHR-OVOL1 system is capable of counteracting skin barrier dysfunction in T-helper type 2-shifted inflammation. This article reviews the dynamic and multifaceted role of AHR in epidermal biology and discusses the potential use of antioxidative phytochemical ligands for AHR in inflammatory skin diseases, such as atopic dermatitis.
Key words: antioxidative phytochemicals; filaggrin; aryl hydrocarbon receptor; nuclear factor-erythroid 2-related factor-2; OVOL1; atopic dermatitis.
Accepted Jul 3, 2018; Epub ahead of print Jul 4, 2018
Acta Derm Venereol 2018; XX: XX–XX.
Corr: Masutaka Furue, Department of Dermatology, Kyushu University, Maidashi 3-1-1, Higashi-ku, Fukuoka 812-8582, Japan. E-mail: furue@dermatol.med.kyushu-u.ac.jp
Aryl hydrocarbon receptor (AHR) is a chemical sensor activated by external and internal ligands. Oxidative AHR ligands induce the production of reactive oxygen species. However, antioxidative phytochemical AHR ligands inhibit ROS generation via activation of nuclear factor-erythroid 2-related factor-2, which is a master switch for antioxidative signalling. In addition, AHR signalling upregulates filaggrin expression via OVOL1 transcription factor and accelerates epidermal terminal differentiation. This AHR-OVOL1 system is capable of counteracting the skin barrier dysfunction in T helper type 2-shifted inflammation. Therefore, antioxidative phytochemicals targeting AHR are potentially beneficial for barrier-disrupted skin diseases, such as atopic dermatitis.
Atopic dermatitis (AD) is characterized by chronic itch, cutaneous inflammation and dry skin with epidermal barrier dysfunction (1–3). Since the discovery of T-helper type 1 (TH1) and TH2 immune regulation by Mosmann et al. (4), the TH2-polarized immune response has been thought to be a cardinal driver in allergic diseases including AD (5–9). This notion has been proven because blockade of TH2-derived interleukin (IL)-4 and IL-13 signalling by a specific anti-IL-4 receptor antibody, dupilumab, successfully improves skin inflammation in patients with AD (10–12). In addition, TH2-derived IL-31 is a potent itching-scratching inducer, and the administration of anti-IL-31 receptor antibody, nemolizumab, improves atopic itching in patients with AD (13–16).
Genome-wide association studies in different ethnicities have revealed at least 19 susceptible genes, including filaggrin (FLG), OVO-like 1 (OVOL1) and IL4/IL13 (17–22). AD exhibits heterogeneous clinical and laboratory manifestations influenced by genetic, environmental and social factors (2, 5, 23–26). However, xerosis or dry skin due to skin barrier disruption is the most frequent clinical sign in AD (23, 25).
Skin barrier maturation is accomplished by sequential and coordinated expression of various terminal differentiation proteins, such as FLG and loricrin (LOR) (27). In accordance, FLG and LOR expression levels have been reported to be reduced in lesioned and non-lesioned skin in AD (28–30). Loss-of-function mutations of FLG have been demonstrated in some patients with AD, ranging from 10% to 50% in the Northern European and Asian AD population (31–34). Ichthyosis vulgaris is also known to be caused by the loss-of function mutation of FLG (35). This may explain why AD is significantly comorbid with ichthyosis vulgaris (25, 31). However, FLG mutations are not found in all patients with AD, and they are less common in Southern Europeans (36) and are even absent in some African countries (37, 38). A humid atmosphere may reduce the contribution of FLG mutations to the onset of AD (39).
Of note, TH2-derived cytokines, IL-4 and IL-13, inhibit FLG and LOR expression (29, 30, 40–42). IL-31 also downregulates FLG and LOR expression (43). Therefore, TH2-polarized inflammatory milieu in AD may be more influential in the downregulation of FLG expression compared with genetic mutations. In line with this notion, topical steroids significantly improve clinical inflammatory signs and normalize transepidermal water loss in lesional AD skin with the upregulation of FLG and LOR expression (44). These improvements are associated with the downregulation of the TH2 (IL-13 and IL-31) signature (44).
Given that the expression levels of FLG and LOR are associated with improvement in AD, strategies to block IL-4-mediated FLG (and LOR) downregulation may be beneficial in treating AD. Although the mechanisms to enhance FLG expression have not been fully understood, recent studies by us and other groups have revealed that aryl hydrocarbon receptor (AHR) signalling plays an essential role in upregulating the expression of FLG and other differentiation-related molecules (29, 30, 45–47). Notably, a plethora of antioxidative phytochemicals work as AHR agonists and restore the IL-4-mediated FLG downregulation (41, 42, 48, 49). These findings have unveiled the underlying mechanisms of how traditionally used antioxidant herbs and phytochemicals work well in maintaining healthy skin and in preventing atopic dry skin. This article focuses on the regulatory mechanisms of AHR/FLG signalling by exogenous antioxidative phytochemicals operating in host epidermal keratinocytes.
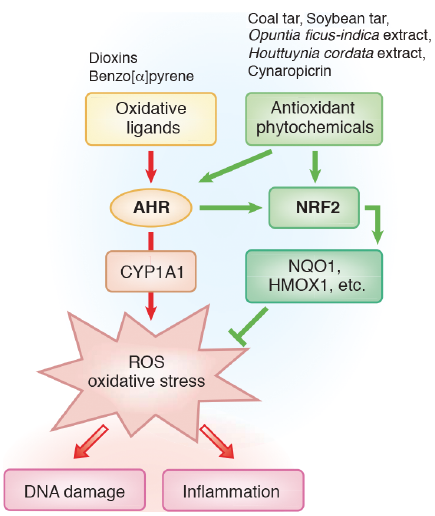
Skin cells, such as keratinocytes, harbour abundant AHR, which exerts multi-functional effects on skin homeostasis and pathology (50, 51). AHR was originally discovered as a cytosolic chemical sensor and transcription factor for halogenated and non-halogenated polycyclic aromatic hydrocarbons, such as 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) and benzo[α]pyrene (50–52). However, in addition to dioxins and benzo[α]pyrene, AHR is a promiscuous receptor, binding with a wide range of affinities to structurally diverse low-molecular-weight chemicals including various phytochemicals (e.g. apigenin, quercetin and cynaropicrin) and tryptophan photoproducts after ultraviolet irradiation (50, 53–56). Most AHR ligands are very hydrophobic; these ligands enter target cells via diffusion and bind to cytosolic AHR. Upon ligand binding, cytosolic AHR undergoes nuclear translocation and binds to its specific DNA recognition site, namely, the xenobiotic-responsive element or dioxin-responsive element, and mediates numerous biological and toxicological effects by inducing the transcription of various AHR-responsive genes, such as cytochrome P4501A1 (CYP1A1) (50, 51, 53) (Fig. 1). In addition to its physiological role in the detoxification of polycyclic aromatic compounds, the activity of the CYP1A1 enzyme can be deleterious, because it generates mutagenic metabolites and reactive oxygen species (ROS) in keratinocytes (50–53). Extensive studies on the function of Ahr using Ahr-deficient mice have demonstrated that Ahr is responsible for most, if not all, of the toxic effects caused by TCDD (50, 51, 53).
Ligands for AHR are divided into at least 2 groups, oxidative and antioxidative ligands (51). Oxidative ligands, such as TCDD and benzo[α]pyrene, induce robust ROS generation in keratinocytes via AHR activation (50, 52). Antioxidative phytochemical ligands, such as coal tar, soybean tar, cynaropicrin, Opuntia ficus-indica extract, Houttuynia cordata extract, Bidens pilosa extract and Galactomyces fermentation, filtrate activate AHR and upregulate CYP1A1 expression, but inhibit ROS generation via the activation of an antioxidant master transcription factor, nuclear factor-erythroid 2-related factor-2 (NRF2) (30, 41, 42, 49, 56–62) (Fig. 1). Oxidative ligands also induce the compensatory activation of NRF2 (63), but the generation of ROS may overwhelm the NRF2-mediated antioxidant activity. Activation of NRF2 induces the transcription of antioxidant enzymes, such as NAD(P)H: quinone oxidoreductase 1 (NQO1) and heme oxygenase-1 (HMOX1), which are the key molecules in achieving antioxidant activity in keratinocytes (58, 59, 62, 64, 65). In line with this notion, ultraviolet B radiation-induced ROS production and sunburn reaction was inhibited via the NRF2/HMOX1 pathway (66, 67).
The bidirectional regulation by AHR in oxidative/antioxidant activity is very functional. For instance, benzo[α]pyrene induces AHR-mediated oxidative stress which is efficiently inhibited by antioxidative phytochemicals via AHR-NRF2 activation (41, 49, 56, 57) (Fig. 2). The antioxidative phytochemical ligands for AHR are also active in counteracting oxidative stress induced by tumour necrosis factor-α (41, 49, 56, 57).
Persistent overactivation of AHR by TCDD and other dioxin-related compounds induces prolonged oxidative stress and may cause chloracne (51, 68, 69). The generation of ROS is also involved in the pathogenesis of contact dermatitis, AD and psoriasis (70–72). Therefore, antioxidative AHR agonists may be beneficial for the treatment of oxidative inflammatory skin diseases. In this context, recent clinical trials have revealed that a natural antioxidative AHR agonist, tapinarof, improves skin lesions of AD and psoriasis in topical use (73–76).
Another intriguing aspect of AHR is its promoting capacity of epidermal terminal differentiation by upregulating FLG, LOR and other differentiation-related molecules (30, 41, 42, 49). These results coincide with the findings that nuclear translocation of AHR is observed in parallel with the terminal differentiation of keratinocytes, and that AHR antagonists have impaired terminal differentiation (77). In parallel, both Ahr-deficient and Ahr-transgenic mice reveal an abnormality in keratinization (78, 79).
Both oxidative and antioxidative AHR ligands induce the coordinated upregulation of FLG, LOR, hornerin and other differentiation-related molecules (41, 42, 47, 49, 77, 80) (Fig. 3). Sustained overactivation of AHR by oxidative TCDD induces exaggerated and accelerated terminal differentiation, which may cause chloracne and hyperkeratosis of the epidermis (47, 80). The oxidative AHR ligands, such as TCDD and benzo[α]pyrene, also induce the keratinocytes to produce proinflammatory cytokines (52, 56, 81).
On the other hand, antioxidative AHR ligands reduce the production of proinflammatory cytokines from keratinocytes (30, 56, 77, 82) and restore the impaired epidermal barrier function in association with FLG upregulation (30, 77, 83, 84). In addition, TH2 cytokine-mediated inhibition of FLG and LOR expression is rescued via AHR activation by antioxidative phytochemicals (29, 30, 41, 42).
A previous study revealed that the FLG gene contains at least 2 xenobiotic-responsive elements where ligated AHR binds (47). Mutations in these sites abrogate AHR-mediated FLG expression (47). In addition to this direct AHR-FLG regulation, we have recently demonstrated a crucial involvement of OVOL1 in the AHR-FLG pathway (29, 48) (Fig. 3). OVOL1 is a transcription factor profoundly related to epithelial differentiation (29, 85, 86) and is highlighted as one of the susceptible genes in AD (19, 22). Abrasion or overexpression of OVOL1 results in the downregulation or upregulation of FLG expression, respectively (29). The activation of AHR by soybean tar Glyteer induces cytoplasmic to nuclear translocation of OVOL1, and this OVOL1 activation results in FLG upregulation (29). IL-4 inhibits FLG expression by blocking the cytoplasmic to nuclear translocation of OVOL1, and the IL-4-induced blockade of OVOL1 translocation is abrogated by AHR activation (29). Thus, AHR ligation rescues IL-4-mediated inhibition of the OVOL1-FLG axis (29). Importantly, the AHR-mediated LOR expression is also mediated by OVOL1 (48). These results stress the importance of AHR signalling in upregulating FLG expression directly or indirectly via OVOL1. Moreover, AHR signalling is effective in counteracting the IL-4-mediated barrier dysfunction in AD (30).
The topical application of some AHR agonists reduces inflammatory skin reactions and restores skin barrier function in mice and humans (76, 84, 87). The beneficial effects of AHR ligation are related, at least in part, to its upregulation capacity of FLG, LOR and other differentiation-related genes in epidermal keratinocytes, as well as anti-inflammatory properties. In addition, some AHR agonists, including various phytochemicals, work as antioxidants via AHR-NRF2 activation. Considering that oxidative stress is exceeded in inflammatory skin conditions, antioxidative AHR agonists are particularly promising in drug development for AD in which TH2-inflammation, barrier disruption and oxidative stress are intermingled.
Funding: This review article was supported by a grant (H27-Shokuhin-Shitei-017) for Research on Food Safety from The Ministry of Health, Labour and Welfare, Japan.
The authors have no conflicts of interest to declare.
- Furue M, Chiba T, Tsuji G, Ulzii D, Kido-Nakahara M, Nakahara T, et al. Atopic dermatitis: immune deviation, barrier dysfunction, IgE autoreactivity and new therapies. Allergol Int 2017; 66: 398–403.
View article Google Scholar - Saeki H, Nakahara T, Tanaka A, Kabashima K, Sugaya M, Murota H, et al. Clinical practice guidelines for the management of atopic dermatitis 2016. J Dermatol 2016; 43: 1117–1145.
View article Google Scholar - Lee SC. A proposal of Reliable Estimation of Atopic Dermatitis in Childhood (REACH) criteria, a novel questionnaire-based diagnostic tool for AD. J Dermatol 2016; 43: 376–384.
View article Google Scholar - Mosmann TR, Cherwinski H, Bond MW, Giedlin MA, Coffman RL. Two types of murine helper T cell clone. I. Definition according to profiles of lymphokine activities and secreted proteins. J Immunol 1986; 136: 2348–2357.
View article Google Scholar - Brunner PM, Guttman-Yassky E, Leung DY. The immunology of atopic dermatitis and its reversibility with broad-spectrum and targeted therapies. J Allergy Clin Immunol 2017; 139: S65–S76.
View article Google Scholar - Czarnowicki T, Esaki H, Gonzalez J, Malajian D, Shemer A, Noda S, et al. Early pediatric atopic dermatitis shows only a cutaneous lymphocyte antigen (CLA)(+) TH2/TH1 cell imbalance, whereas adults acquire CLA(+) TH22/TC22 cell subsets. J Allergy Clin Immunol 2015; 136: 941–951.
View article Google Scholar - Esaki H, Brunner PM, Renert-Yuval Y, Czarnowicki T, Huynh T, Tran G, et al. Early-onset pediatric atopic dermatitis is T(H)2 but also T(H)17 polarized in skin. J Allergy Clin Immunol 2016; 138: 1639–1651.
View article Google Scholar - Esaki H, Takeuchi S, Furusyo N, Yamamura K, Hayashida S, Tsuji G, et al. Levels of immunoglobulin E specific to the major food allergen and chemokine (C-C motif) ligand (CCL)17/thymus and activation regulated chemokine and CCL22/macrophage-derived chemokine in infantile atopic dermatitis on Ishigaki Island. J Dermatol 2016; 43: 1278–1282.
View article Google Scholar - Miyano K, Matsushita S, Tsuchida T, Nakamura K. Inhibitory effect of a histamine 4 receptor antagonist on CCL17 and CCL22 production by monocyte-derived Langerhans cells in patients with atopic dermatitis. J Dermatol 2016; 43: 1024–1029.
View article Google Scholar - Blauvelt A, de Bruin-Weller M, Gooderham M, Cather JC, Weisman J, Pariser D, et al. Long-term management of moderate-to-severe atopic dermatitis with dupilumab and concomitant topical corticosteroids (LIBERTY AD CHRONOS): a 1-year, randomised, double-blinded, placebo-controlled, phase 3 trial. Lancet 2017; 389: 2287–2303.
View article Google Scholar - de Bruin-Weller M, Thaçi D, Smith CH, Reich K, Cork M, Radin A, et al. Dupilumab with concomitant topical corticosteroids in adult patients with atopic dermatitis who are not adequately controlled with or are intolerant to ciclosporin A, or when this treatment is medically inadvisable: a placebo-controlled, randomized phase 3 clinical trial (LIBERTY AD CAFÉ). Br J Dermatol 2018; 178: 1083–1101.
View article Google Scholar - Simpson EL, Bieber T, Guttman-Yassky E, Beck LA, Blauvelt A, Cork MJ, et al. Two phase 3 trials of dupilumab versus placebo in atopic dermatitis. N Engl J Med 2016; 375: 2335–2348.
View article Google Scholar - Furue M, Yamamura K, Kido-Nakahara M, Nakahara T, Fukui Y. Emerging role of interleukin-31 and interleukin-31 receptor in pruritus in atopic dermatitis. Allergy 2018; 73: 29–36.
View article Google Scholar - Ruzicka T, Hanifin JM, Furue M, Pulka G, Mlynarczyk I, Wollenberg A, et al. Anti-interleukin-31 receptor A antibody for atopic dermatitis. N Engl J Med 2017; 376: 826–835.
View article Google Scholar - Nemoto O, Furue M, Nakagawa H, Shiramoto M, Hanada R, Matsuki S, et al. The first trial of CIM331, a humanized antihuman interleukin-31 receptor A antibody, in healthy volunteers and patients with atopic dermatitis to evaluate safety, tolerability and pharmacokinetics of a single dose in a randomized, double-blind, placebo-controlled study. Br J Dermatol 2016; 174: 296–304.
View article Google Scholar - Yamamura K, Uruno T, Shiraishi A, Tanaka Y, Ushijima M, Nakahara T, et al. The transcription factor EPAS1 links DOCK8 deficiency to atopic skin inflammation via IL-31 induction. Nat Commun 2017; 8: 13946.
View article Google Scholar - Ellinghaus D, Baurecht H, Esparza-Gordillo J, Rodríguez E, Matanovic A, Marenholz I, et al. High-density genotyping study identifies four new susceptibility loci for atopic dermatitis. Nat Genet 2013; 45: 808–812.
View article Google Scholar - Esparza-Gordillo J, Weidinger S, Fölster-Holst R, Bauerfeind A, Ruschendorf F, Patone G, et al. A common variant on chromosome 11q13 is associated with atopic dermatitis. Nat Genet 2009; 41: 596–601.
View article Google Scholar - Hirota T, Takahashi A, Kubo M, Tsunoda T, Tomita K, Sakashita M, et al. Genome-wide association study identifies eight new susceptibility loci for atopic dermatitis in the Japanese population. Nat Genet 2012; 44: 1222–1226.
View article Google Scholar - Paternoster L, Standl M, Chen CM, Ramasamy A, Bønnelykke K, Duijts L, et al. Meta-analysis of genome-wide association studies identifies three new risk loci for atopic dermatitis. Nat Genet 2011; 44: 187–192.
View article Google Scholar - Sun LD, Xiao FL, Li Y, Zhou WM, Tang HY, Tang XF, et al. Genome-wide association study identifies two new susceptibility loci for atopic dermatitis in the Chinese Han population. Nat Genet 2011; 43: 690–694.
View article Google Scholar - Tamari M, Hirota T. Genome-wide association studies of atopic dermatitis. J Dermatol 2014; 41: 213–220.
View article Google Scholar - Chu H, Shin JU, Park CO, Lee H, Lee J, Lee KH. Clinical diversity of atopic dermatitis: a review of 5,000 patients at a single institute. Allergy Asthma Immunol Res 2017; 9: 158–168.
View article Google Scholar - Furue M, Chiba T, Takeuchi S. Current status of atopic dermatitis in Japan. Asia Pac Allergy 2011; 1: 64–72.
View article Google Scholar - Hanifin JM, Rajka G. Diagnostic features of atopic eczema. Acta Derm Venereol Suppl 1980; 92: 44–47.
View article Google Scholar - Murota H, Takeuchi S, Sugaya M, Tanioka M, Onozuka D, Hagihara A, et al. Characterization of socioeconomic status of Japanese patients with atopic dermatitis showing poor medical adherence and reasons for drug discontinuation. J Dermatol Sci 2015; 79: 279–287.
View article Google Scholar - Kypriotou M, Huber M, Hohl D. The human epidermal differentiation complex: cornified envelope precursors, S100 proteins and the ‘fused genes’ family. Exp Dermatol 2012; 21: 643–649.
View article Google Scholar - Seguchi T, Cui CY, Kusuda S, Takahashi M, Aisu K, Tezuka T. Decreased expression of filaggrin in atopic skin. Arch Dermatol Res 1996; 288: 442–446.
View article Google Scholar - Tsuji G, Hashimoto-Hachiya A, Kiyomatsu-Oda M, Takemura M, Ohno F, Ito T, et al. Aryl hydrocarbon receptor activation restores filaggrin expression via OVOL1 in atopic dermatitis. Cell Death Dis 2017; 8: e2931.
View article Google Scholar - van den Bogaard EH, Bergboer JG, Vonk-Bergers M, van Vlijmen-Willems IM, Hato SV, van der Valk PG, et al. Coal tar induces AHR-dependent skin barrier repair in atopic dermatitis. J Clin Invest 2013; 123: 917–927.
View article Google Scholar - McLean WH. Filaggrin failure – from ichthyosis vulgaris to atopic eczema and beyond. Br J Dermatol 2016; 175 Suppl 2: 4–7.
View article Google Scholar - Nomura T, Sandilands A, Akiyama M, Liao H, Evans AT, Sakai K, et al. Unique mutations in the filaggrin gene in Japanese patients with ichthyosis vulgaris and atopic dermatitis. J Allergy Clin Immunol 2007; 119: 434–440.
View article Google Scholar - Palmer CN, Irvine AD, Terron-Kwiatkowski A, Zhao Y, Liao H, Lee SP, et al. Common loss-of-function variants of the epidermal barrier protein filaggrin are a major predisposing factor for atopic dermatitis. Nat Genet 2006; 38: 441–446.
View article Google Scholar - Sandilands A, Terron-Kwiatkowski A, Hull PR, O’Regan GM, Clayton TH, Watson RM, et al. Comprehensive analysis of the gene encoding filaggrin uncovers prevalent and rare mutations in ichthyosis vulgaris and atopic eczema. Nat Genet 2007; 39: 650–654.
View article Google Scholar - Smith FJ, Irvine AD, Terron-Kwiatkowski A, Sandilands A, Campbell LE, Zhao Y, et al. Loss-of-function mutations in the gene encoding filaggrin cause ichthyosis vulgaris. Nat Genet 2006; 38: 337–342.
View article Google Scholar - Cascella R, Foti Cuzzola V, Lepre T, Galli E, Moschese V, Chini L, et al. Full sequencing of the FLG gene in Italian patients with atopic eczema: evidence of new mutations, but lack of an association. J Invest Dermatol 2011; 131: 982–984.
View article Google Scholar - Thawer-Esmail F, Jakasa I, Todd G, Wen Y, Brown SJ, Kroboth K, et al. South African amaXhosa patients with atopic dermatitis have decreased levels of filaggrin breakdown products but no loss-of-function mutations in filaggrin. J Allergy Clin Immunol 2014; 133: 280–282.
View article Google Scholar - Winge MC, Bilcha KD, Liedén A, Shibeshi D, Sandilands A, Wahlgren CF, et al. Novel filaggrin mutation but no other loss-of-function variants found in Ethiopian patients with atopic dermatitis. Br J Dermatol 2011; 165: 1074–1080.
View article Google Scholar - Sasaki T, Furusyo N, Shiohama A, Takeuchi S, Nakahara T, Uchi H, et al. Filaggrin loss-of-function mutations are not a predisposing factor for atopic dermatitis in an Ishigaki Island under subtropical climate. J Dermatol Sci 2014; 76: 10–15.
View article Google Scholar - Howell MD, Kim BE, Gao P, Grant AV, Boguniewicz M, Debenedetto A, et al. Cytokine modulation of atopic dermatitis filaggrin skin expression. J Allergy Clin Immunol 2007; 120: 150–155.
View article Google Scholar - Takei K, Mitoma C, Hashimoto-Hachiya A, Uchi H, Takahara M, Tsuji G, et al. Antioxidant soybean tar Glyteer rescues T-helper-mediated downregulation of filaggrin expression via aryl hydrocarbon receptor. J Dermatol 2015; 42: 171–180.
View article Google Scholar - Takei K, Mitoma C, Hashimoto-Hachiya A, Takahara M, Tsuji G, Nakahara T, et al. Galactomyces fermentation filtrate prevents T helper 2-mediated reduction of filaggrin in an aryl hydrocarbon receptor-dependent manner. Clin Exp Dermatol 2015; 40: 786–793.
View article Google Scholar - Cornelissen C, Marquardt Y, Czaja K, Wenzel J, Frank J, Lüscher-Firzlaff J, et al. IL-31 regulates differentiation and filaggrin expression in human organotypic skin models. J Allergy Clin Immunol 2012; 129: 426–433.
View article Google Scholar - Guttman-Yassky E, Ungar B, Malik K, Dickstein D, Suprun M, Estrada YD, et al. Molecular signatures order the potency of topically applied anti-inflammatory drugs in patients with atopic dermatitis. J Allergy Clin Immunol 2017; 140: 1032–1042.
View article Google Scholar - Furue M, Tsuji G, Mitoma C, Nakahara T, Chiba T, Morino-Koga S, et al. Gene regulation of filaggrin and other skin barrier proteins via aryl hydrocarbon receptor. J Dermatol Sci 2015; 80: 83–88.
View article Google Scholar - Sutter CH, Yin H, Li Y, Mammen JS, Bodreddigari S, Stevens G, et al. EGF receptor signaling blocks aryl hydrocarbon receptor-mediated transcription and cell differentiation in human epidermal keratinocytes. Proc Natl Acad Sci U S A 2009; 106: 4266–4271.
View article Google Scholar - Sutter CH, Bodreddigari S, Campion C, Wible RS, Sutter TR. 2,3,7,8-Tetrachlorodibenzo-p-dioxin increases the expression of genes in the human epidermal differentiation complex and accelerates epidermal barrier formation. Toxicol Sci 2011; 124: 128–137.
View article Google Scholar - Hirano A, Goto M, Mitsui T, Hashimoto-Hachiya A, Tsuji G, Furue M. Antioxidant Artemisia princeps extract enhances the expression of filaggrin and loricrin via the AHR/OVOL1 pathway. Int J Mol Sci 2017; 18: E1948.
View article Google Scholar - Nakahara T, Mitoma C, Hashimoto-Hachiya A, Takahara M, Tsuji G, Uchi H, et al. Antioxidant Opuntia ficus-indica extract activates AHR-NRF2 signaling and upregulates filaggrin and loricrin expression in human keratinocytes. J Med Food 2015; 18: 1143–1149.
View article Google Scholar - Esser C, Bargen I, Weighardt H, Haarmann-Stemmann T, Krutmann J. Functions of the aryl hydrocarbon receptor in the skin. Semin Immunopathol 2013; 35: 677–691.
View article Google Scholar - Furue M, Takahara M, Nakahara T, Uchi H. Role of AhR/ARNT system in skin homeostasis. Arch Dermatol Res 2014; 306: 769–779.
View article Google Scholar - Tsuji G, Takahara M, Uchi H, Takeuchi S, Mitoma C, Moroi Y, et al. An environmental contaminant, benzo(a)pyrene, induces oxidative stress-mediated interleukin-8 production in human keratinocytes via the aryl hydrocarbon receptor signaling pathway. J Dermatol Sci 2011; 62: 42–49.
View article Google Scholar - Denison MS, Soshilov AA, He G, DeGroot DE, Zhao B. Exactly the same but different: promiscuity and diversity in the molecular mechanisms of action of the aryl hydrocarbon (dioxin) receptor. Toxicol Sci 2011; 124: 1–22.
View article Google Scholar - Fritsche E, Schäfer C, Calles C, Bernsmann T, Bernshausen T, Wurm M, et al. Lightening up the UV response by identification of the aryl hydrocarbon receptor as a cytoplasmatic target for ultraviolet B radiation. Proc Natl Acad Sci U S A 2007; 104: 8851–8856.
View article Google Scholar - Nguyen LP, Bradfield CA. The search for endogenous activators of the aryl hydrocarbon receptor. Chem Res Toxicol 2008; 21: 102–116.
View article Google Scholar - Takei K, Hashimoto-Hachiya A, Takahara M, Tsuji G, Nakahara T, Furue M. Cynaropicrin attenuates UVB-induced oxidative stress via the AhR-Nrf2-Nqo1 pathway. Toxicol Lett 2015; 234: 74–80.
View article Google Scholar - Doi K, Mitoma C, Nakahara T, Uchi H, Hashimoto-Hachiya A, Takahara M, et al. Antioxidant Houttuynia cordata extract upregulates filaggrin expression in an aryl hydrocarbon-dependent manner. Fukuoka Igaku Zasshi 2014; 105: 205–213.
View article Google Scholar - G?gotek A, Skrzydlewska E. The role of transcription factor Nrf2 in skin cells metabolism. Arch Dermatol Res 2015; 307: 385–396.
View article Google Scholar - Haarmann-Stemmann T, Abel J, Fritsche E, Krutmann J. The AhR-Nrf2 pathway in keratinocytes: on the road to chemoprevention? J Invest Dermatol 2012; 132: 7–9.
View article Google Scholar - Kohda F, Takahara M, Hachiya A, Takei K, Tsuji G, Yamamura K, et al. Decrease of reactive oxygen species and reciprocal increase of nitric oxide in human dermal endothelial cells by Bidens pilosa extract: a possible explanation of its beneficial effect on livedo vasculopathy. J Dermatol Sci 2013; 72: 75–77.
View article Google Scholar - Qin S, Hou DX. Multiple regulations of Keap1/Nrf2 system by dietary phytochemicals. Mol Nutr Food Res 2016; 60: 1731–1755.
View article Google Scholar - Tsuji G, Takahara M, Uchi H, Matsuda T, Chiba T, Takeuchi S, et al. Identification of ketoconazole as an AhR-Nrf2 activator in cultured human keratinocytes: the basis of its anti-inflammatory effect. J Invest Dermatol 2012; 132: 59–68.
View article Google Scholar - Wang L, He X, Szklarz GD, Bi Y, Rojanasakul Y, Ma Q. The aryl hydrocarbon receptor interacts with nuclear factor erythroid 2-related factor 2 to mediate induction of NAD(P)H: quinoneoxidoreductase 1 by 2,3,7,8-tetrachlorodibenzo-p-dioxin. Arch Biochem Biophys 2013; 537: 31–38.
View article Google Scholar - Tsuji G, Hashimoto-Hachiya A, Takemura M, Kanemaru T, Ichihashi M, Furue M. Palladium and platinum nanoparticles activate AHR and NRF2 in human keratinocytes-implications in vitiligo therapy. J Invest Dermatol 2017; 137: 1582–1586.
View article Google Scholar - Uchi H, Yasumatsu M, Morino-Koga S, Mitoma C, Furue M. Inhibition of aryl hydrocarbon receptor signaling and induction of NRF2-mediated antioxidant activity by cinnamaldehyde in human keratinocytes. J Dermatol Sci 2017; 85: 36–43.
View article Google Scholar - Tigges J, Haarmann-Stemmann T, Vogel CFA, Grindel A, Hübenthal U, Brenden H, et al. The new aryl hydrocarbon receptor antagonist E/Z-2-benzylindene-5,6-dimethoxy-3,3-dimethylindan-1-one protects against UVB-induced signal transduction. J Invest Dermatol 2014; 134: 556–559.
View article Google Scholar - Wu Z, Uchi H, Morino-Koga S, Shi W, Furue M. Z-ligustilide ameliorated ultraviolet B-induced oxidative stress and inflammatory cytokine production in human keratinocytes through upregulation of Nrf2/HO-1 and suppression of NF-?B pathway. Exp Dermatol 2015; 24: 703–708.
View article Google Scholar - Mitoma C, Mine Y, Utani A, Imafuku S, Muto M, Akimoto T, et al. Current skin symptoms of Yusho patients exposed to high levels of 2,3,4,7,8-pentachlorinated dibenzofuran and polychlorinated biphenyls in 1968. Chemosphere 2015; 137: 45–51.
View article Google Scholar - Mitoma C, Uchi H, Tsukimori K, Yamada H, Akahane M, Imamura T, et al. Yusho and its latest findings – a review in studies conducted by the Yusho Group. Environ Int 2015; 82: 41–48.
View article Google Scholar - Eto H, Tsuji G, Chiba T, Furue M, Hyodo F. Non-invasive evaluation of atopic dermatitis based on redox status using in vivo dynamic nuclear polarization magnetic resonance imaging. Free Radic Biol Med 2016; 103: 209–215.
View article Google Scholar - Lee YJ, Bae JH, Kang SG, Cho SW, Chun DI, Nam SM, et al. Pro-oxidant status and Nrf2 levels in psoriasis vulgaris skin tissues and dimethyl fumarate-treated HaCaT cells. Arch Pharm Res 2017; 40: 1105–1116.
View article Google Scholar - Yasukawa S, Miyazaki Y, Yoshii C, Nakaya M, Ozaki N, Toda S, et al. An ITAM-Syk-CARD9 signalling axis triggers contact hypersensitivity by stimulating IL-1 production in dendritic cells. Nat Commun 2014; 5: 3755.
View article Google Scholar - Bissonnette R, Poulin Y, Zhou Y, Tan J, Hong HC, Webster J, et al. Efficacy and safety of topical WBI-1001 in patients with mild to severe atopic dermatitis: results from a 12-week, multicentre, randomized, placebo-controlled double-blind trial. Br J Dermatol 2012; 166: 853–860.
View article Google Scholar - Bissonnette R, Bolduc C, Maari C, Nigen S, Webster JM, Tang L, et al. Efficacy and safety of topical WBI-1001 in patients with mild to moderate psoriasis: results from a randomized double-blind placebo-controlled, phase II trial. J Eur Acad Dermatol Venereol 2012; 26: 1516–1521.
View article Google Scholar - Johnston A. Resolving inflammation by targeting an ancient innate immune sensor with a bacterial metabolite. J Invest Dermatol 2017; 137: 2050–2052.
View article Google Scholar - Smith SH, Jayawickreme C, Rickard DJ, Nicodeme E, Bui T, Simmons C, et al. Tapinarof Is a natural AhR agonist that resolves skin inflammation in mice and humans. J Invest Dermatol 2017; 137: 2110–2119.
View article Google Scholar - van den Bogaard EH, Podolsky MA, Smits JP, Cui X, John C, Gowda K, et al. Genetic and pharmacological analysis identifies a physiological role for the AHR in epidermal differentiation. J Invest Dermatol 2015; 135: 1320–1328.
View article Google Scholar - Fernandez-Salguero PM, Ward JM, Sundberg JP, Gonzalez FJ. Lesions of aryl-hydrocarbon receptor-deficient mice. Vet Pathol 1997; 34: 605–614.
View article Google Scholar - Tauchi M, Hida A, Negishi T, Katsuoka F, Noda S, Mimura J, et al. Constitutive expression of aryl hydrocarbon receptor in keratinocytes causes inflammatory skin lesions. Mol Cell Biol 2005; 25: 9360–9368.
View article Google Scholar - Kennedy LH, Sutter CH, Leon Carrion S, Tran QT, Bodreddigari S, Kensicki E, et al. 2,3,7,8-Tetrachlorodibenzo-p-dioxin-mediated production of reactive oxygen species is an essential step in the mechanism of action to accelerate human keratinocyte differentiation. Toxicol Sci 2013; 132: 235–249.
View article Google Scholar - Kim HO, Kim JH, Chung BY, Choi MG, Park CW. Increased expression of the aryl hydrocarbon receptor in patients with chronic inflammatory skin diseases. Exp Dermatol 2014; 23: 278–281.
View article Google Scholar - Amagai Y, Katsuta C, Nomura Y, Oida K, Matsuda K, Jang H, et al. Amelioration of atopic-like skin conditions in NC/Tnd mice by topical application with distilled Alpinia intermedia Gagnep extracts. J Dermatol 2017; 44: 1238–1247.
View article Google Scholar - Haas K, Weighardt H, Deenen R, Köhrer K, Clausen B, Zahner S, et al. Aryl Hydrocarbon receptor in keratinocytes is essential for murine skin barrier integrity. J Invest Dermatol 2016; 136: 2260–2269.
View article Google Scholar - Hou M, Sun R, Hupe M, Kim PL, Park K, Crumrine D, et al. Topical apigenin improves epidermal permeability barrier homoeostasis in normal murine skin by divergent mechanisms. Exp Dermatol 2013; 22: 210–215.
View article Google Scholar - Lee B, Villarreal-Ponce A, Fallahi M, Ovadia J, Sun P, Yu QC, et al. Transcriptional mechanisms link epithelial plasticity to adhesion and differentiation of epidermal progenitor cells. Dev Cell 2014; 29: 47–58.
View article Google Scholar - Nair M, Teng A, Bilanchone V, Agrawal A, Li B, Dai X. Ovol1 regulates the growth arrest of embryonic epidermal progenitor cells and represses c-myc transcription. J Cell Biol 2006; 173: 253–264.
View article Google Scholar - Di Meglio P, Duarte JH, Ahlfors H, Owens ND, Li Y, Villanova F, et al. Activation of the aryl hydrocarbon receptor dampens the severity of inflammatory skin conditions. Immunity 2014; 40: 989–1001.
View article Google Scholar





 Click to show fullsize
Click to show fullsize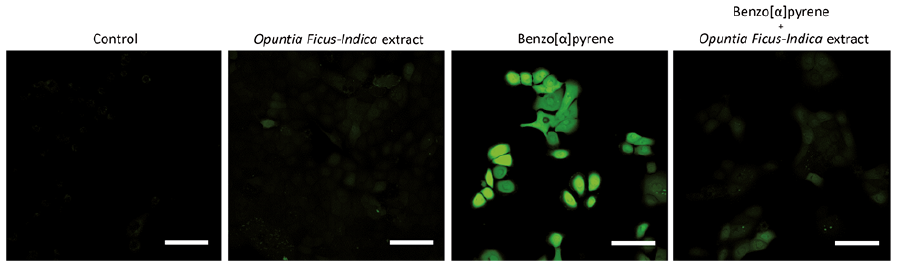 Click to show fullsize
Click to show fullsize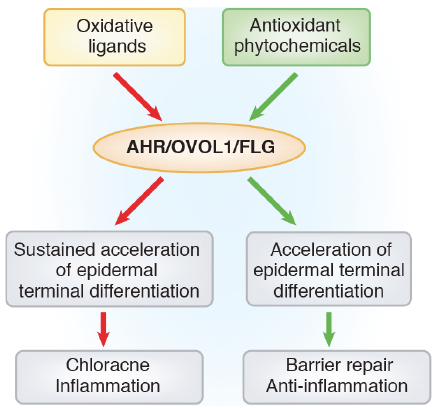 Click to show fullsize
Click to show fullsize