


1Department of Dermatology, Shandong Provincial Hospital for Skin Disease, Shandong University, 2Shandong Provincial Institute of Dermatology and Venereology, Shandong Academy of Medical Sciences, Jinan, Shandong, 3Shandong Provincial Key Laboratory for Dermatovenereology, 4School of Medicine and Life Science, University of Jinan-Shandong Academy of Medical Sciences, Jinan, Shandong, and 5Binzhou Medical University, Yantai, Shandong, China
Genome-wide association studies have recently identified a number of non-major histocompatibility complex regions associated with psoriatic arthritis. However, data on Chinese patients with psoriatic arthritis and the differences between psoriatic arthritis and cutaneous psoriasis are limited. This study genotyped 12 single nucleotide polymorphisms in 379 patients with psoriatic arthritis, 376 with cutaneous psoriasis, and 760 healthy controls using Sequenom’s Mass ARRAY system. The aim of the study was to expand the database for psoriatic arthritis and cutaneous psoriasis, and develop a genetic prediction system for the early diagnosis of psoriatic arthritis in the Chinese population. One variant in NFKBIA, rs12883343, had a significantly different association with psoriatic arthritis than with cutaneous psoriasis (p = 4.93×10–10, odds ratio 2.371). This suggests that there are differences in the pathogenesis of psoriatic arthritis and cutaneous psoriasis.
Key words: psoriatic arthritis; cutaneous psoriasis; NFKBIA.
Accepted Aug 31, 2018; Epub ahead of print Sep 3, 2018
Acta Derm Venereol 2018; XX: XX–XX.
Corr: Furen Zhang and Qing Yang, Department of Dermatology and Venereology, Shandong Provincial Institute of Dermatology and Venereology, Shandong Academy of Medical Science, 27397 Jingshi Lu, Shandong Province, Jinan 250022, China. E-mails: zhangfuren@hotmail.com; yangqing79@126.com
A number of novel psoriatic arthritis genetic susceptibility loci have been identified recently, but data on their association with Chinese psoriatic arthritis patients are limited. In addition, the data of differences between psoriatic arthritis and cutaneous psoriasis are lacking. We performed a genetic study in a Chinese population to expand the database for psoriatic arthritis and cutaneous psoriasis and to develop a genetic prediction model for early diagnosis of psoriatic arthritis.
Psoriatic arthritis (PsA) is an inflammatory arthritis associated with psoriasis, characterized by seronegative rheumatoid factor. Among patients with psoriasis, the prevalence of PsA varies from 6% in the USA to 42% in South Africa (1), while published figures indicate lower prevalence rates in Asian countries (2), such as Korea (9%) (3) and Japan (1%) (4). In China, the prevalence of PsA has been estimated as 5.8% (5) based on a large cross-sectional observational study. Although the exact mechanism of PsA is unclear, host genetics, immunological, and environmental factors are thought to play a role (6).
With the development of genome-wide association studies (GWAS), 4 PsA GWAS have been published to date. Together with candidate loci analyses and Immunochip array studies, 15 non-major histocompatibility complex (non-MHC) regions associated with PsA have shown statistical significance (p ≤ 5×10–8). These include IL-12B (7–10), IL-23R (7, 10), TNIP1 (7, 8, 10), TRAF3IP2 (7, 9, 11, 12), CSF2 (7), FBXL19 (13), REL (7, 14), RUNX3 (15), TYK2 (7, 10), NOS2 (16), PTPN22 (16), IFNLR1 (7), IFIH1 (7), NFKBIA (7) and STAT2 (10). Current genetic studies have suggested that key pathways in the pathogenesis of PsA include the NFκB and IFN signalling pathway, and adaptive immune responses involving CD8+T cells and CD4+T helper (Th) 17-cell signalling (17–20).
Clinically, psoriasis vulgaris (PsV) is characterized by sharply demarcated, scaly, erythematous plaques. As with PsV, PsA has the common manifestation of skin lesions, but additionally manifests with peripheral arthritis, enthesitis, dactylitis, uveitis, and spondylitis (20). GWAS and candidate loci analyses have revealed differences in their genetic architecture (7). Significant differences in the strength of association between psoriasis and the MHC have been observed. It has been reported that the human leukocyte-associated antigen (HLA)-Cw*0602 is strongly associated with PsV (21), while HLA-B*27 plays an important role in PsA (22). Winchester et al. (22) hypothesized that the MHC molecules encoded by HLA-C*0602 presented skin-specific self-peptides to T cells, whereas B*27 molecules are mediators for the primary response to a cutaneous and joint antigen. Currently, there are 13 non-MHC regions with a nominally significant (p < 0.05) difference in their association for these 2 forms of psoriasis (7). However, limited data are available for China. Yang et al. (23) reported single nucleotide polymorphisms (SNPs) at IL12B and ZNF816A, with a nominal p-value (p < 0.05) between PsA and PsV.
To address the need to expand the database for PsA and cutaneous psoriasis (PsC), 15 non-MHC PsA susceptibility loci were tested in a Chinese cohort involving 379 patients with PsA, 376 with PsC and 760 unaffected control individuals. The aim of the study was to develop a genetic prediction model for early diagnosis of PsA in the Chinese population.
A total of 755 patients with psoriasis (379 patients with PsA and 376 with PsC) were included. All patients were recruited from the Shandong Provincial Hospital for Skin Disease. Patients were diagnosed by both a dermatologist and rheumatologist. All PsA case subjects met the Classification for Psoriatic Arthritis (CASPAR) criteria (24). PsC was defined as an individual having been a PsV patient for 10 or more years without developing any signs of PsA (7). All patients were resident in Shandong province and had no reported genetic relationship. Healthy controls in this study were all Han Chinese from the same region, with no history of psoriasis or autoimmune diseases. Clinical information was collected from the subjects through a full clinical check-up, while additional information was obtained through a questionnaire. All participants provided written informed consent. The study was approved by the ethics committee of Shandong Provincial Institute of Dermatology and Venereology and was conducted according to the principles of the Declaration of Helsinki.
Fifteen SNP markers were selected from 4 GWASs and candidate-gene association studies (7–16). SNPs were selected based on the following criteria: (i) with the exception of rs9321623 at TNFAIP3, each SNP selected from the GWAS showed that statistically significant differences (p ≤ 5×10–8) exist between PsA and control samples; (ii) all rare SNPs in Asian populations were excluded (minor allele frequency (MAF), obtained from 1,000 Genomes Phase 1 Asian data, was < 0.05); (iii) SNPs were independent of each other (linkage disequilibrium (LD) r2 ≤ 0.5); and (iv) SNPs were associated with coding variants or potential regulatory elements with a score ≤ 5 in the Regulomedb or < 5 in the Encode database.
Genotyping analysis of all the samples was conducted using Sequenom’s Mass ARRAY system. DNA was amplified through multiplex PCR using 15 ng genomic DNA. PCR products were then used for locus-specific single-based extension reactions, desalted, and transferred to the SpectroCHIP array. Allele detection was performed using MALDI-TOF mass spectrometry. The mass spectrograms were analysed using Sequenom Mass ARRAY TYPER software. SNPs with a call rate < 90%, or a deviation from Hardy–Weinberg equilibrium (p < 0.01) in the control subjects were excluded from further analysis. Of the 15 SNPs subjected to the validation study, 3 (rs9321623, rs34725611, rs35251378) failed in the genotyping analysis and were excluded from further analysis.
Associations were tested based on a logistic model grouped by PsA patients vs. controls, PsC patients vs. controls, and PsA patients vs. PsC patients using PLINK v 1.07 (25). Logistic regression for each SNP was performed by adjusting for sex. In order to correct the number of loci studied, p < 0.004 was accepted by a Bonferroni correction of 12.
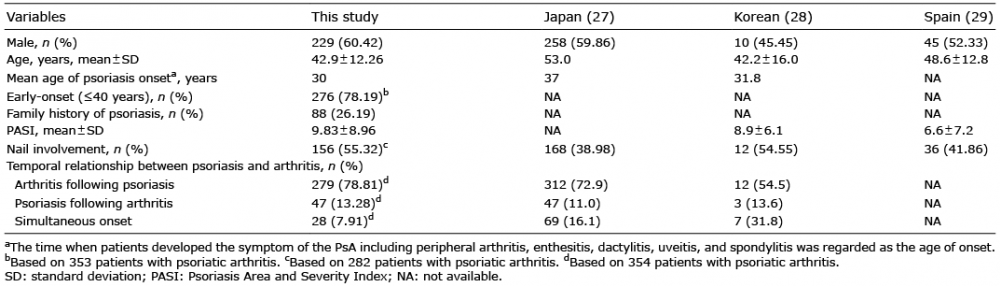
The demographic and clinical characteristics of patients with PsA are shown in Table I. The mean age at onset of patients with PsA was 42.9 years. Patients with PsA had a higher rate of severe psoriasis with a Psoriasis Area and Severity Index (PASI) score 9.83 higher than those without PsA. As demonstrated in previous reports, nail involvement was more common in patients with PsA (55.32%) than in those without PsA (21.0%) (5). Among patients with PsA, arthritis preceded psoriasis in 13.28% of cases. The temporal relationship between psoriasis and arthritis is shown in Table I. According to the Moll and Wright classification criteria (26), the manifestation of PsA included spondylitis in 15.22% of patients, oligoarthritis in 71.64%, polyarthritis in 8.66%, predominant distal interphalangeal joint (DIP) arthritis in 2.99%, and arthritis mutilans in 1.49% (Table II).
Table I. Clinical characteristics for patients with psoriatic arthritis
Table II. Manifestation pattern of patients with psoriatic arthritisa
Compared with Asian and European data (Table I), there is not a great difference between our results and those of previous studies conducted in other Asia and Western countries. This study, as well as previous studies, suggests that PsA affects men more often than women. Psoriasis presented prior to PsA in >50% of patients. The mean age at onset of PsA was approximately 40–50 years and the mean age of psoriasis onset in PsA cohorts was 30–40 years. In addition, nail involvement was quite common.
Based on the results of previous publications, 15 SNPs that were above a genome-wide significance level were selected for subsequent validation. After quality control, one patient with PsA and 11 patients with PsC were removed as the call rate was < 90%. Finally, 378 PsA, 365 PsC and 760 controls samples were available for validation analysis. The characteristics of the samples are shown in Table III. Three SNPs failed to meet the inclusion criteria during assay design.
Table III. Summary information for psoriatic arthritis (PsA) and cutaneous psoriasis (PsC) cases and controls
Firstly, the association of the SNPs with a state of psoriasis was evaluated by comparing PsA with control samples. By comparing all PsA cases against all control samples, disease association was replicated in 5 of the 12 SNPs tested. However, all SNP associations failed Bonferroni correction in the PsA cohort (Table IV). Mapping to IL12B, the strongest association was identified at a genome-wide significance level for rs2082412 (p = 4.32 × 10–3, odds ratio (OR) 0.766), which corroborates with data reported by Yang et al. (23). Four other SNPs were found to be associated with PsA in the Chinese population in this study. This included: rs 4921485 at IL12B (p = 4.50 × 10–3, OR 0.767); rs12924903 at FBXL19 (p = 7.48 × 10–3, OR 1.540); rs 4655683 at IL23R (p = 1.11 × 10–2, OR 1.254); and rs12044149 at IL23R (p = 1.24 × 10–2, OR 1.264). Within the set of positively replicated loci, IFNLR1, RUNX3, IFIH1, TRAF3IP2, NFKBIA, and NOS2 had not been tested in an independent replication sample, unlike the previously studies.
Table IV. Results for previously reported single nucleotide polymorphisms (SNPs) comparative strength of association of psoriatic arthritis (PsA) vs. cutaneous psoriasis (PsC)
Next, we evaluated the disease association of the 12 SNPs by comparing the PsC and control samples. Two SNPs were associated with PsC. The rs12883343 SNP (NFKBIA) was significantly associated with PsC (p = 1.26 × 10–8, OR 0.486), while the rs2082412 SNP (IL12B) showed borderline evidence for PsC association at a nominal p-value threshold of 0.05 (p = 1.60 × 10–2, OR 0.798). Finally, we compared the genotypic frequencies of these SNPs across PsA and PsC cohorts. Our study demonstrated that the NFKBIA rs12883343 variant exhibited a significantly different association with PsA compared with PsC (p = 4.93 × 10–10, OR 2.371). Another variant at IL12B, which has not been described previously, rs4921485, indicated borderline evidence for disease association (p = 2.84 × 10–2, OR 0.785). The rs4921485 showed evidence of an association between axial and peripheral PsA subgroups (p = 7.8 × 10–3, OR 1.548), but this failed the Bonferroni correction. We also confirmed the previous findings that the non-MHC variant rs 4655683 at IL23R is associated with disease (p = 2.56 × 10–2, OR 1.273).
PsA has a complex genetic background. Recently, a large number of loci that are confidently associated with the risk of developing this disease were identified in large GWAS population cohorts (7). However, data representative of the Chinese population are limited. This study provides significant evidence for the genetic associations of PsA and PsC in the Chinese population. A genetic association between PsA and PsC was also identified. We furthermore found that the NFKBIA locus was involved in PsC susceptibility. Finally, we also discovered and describe a previously unknown, independent variant that is differentially associated with PsA compared with PsC.
In 78.8% patients with PsA in our study skin manifestations preceded joint symptoms. Therefore, it is vital to not only identify new therapeutic targets, but also pinpoint disease-specific genetic risk factors in order to predict which psoriasis patients are at high risk of developing PsA. We discovered a variant in the NFKBIA region (rs12883343), which exhibited a significantly different association with PsA and PsC. NFKBIA as well as NKBI and RelA are the components of the nuclear factor-κB (NF-κB) complex. NFKBIA encodes I-kappa-B (NFKBIα), which blocks the NF-κB nuclear localizing sequence, thereby inactivating the complex in the cytoplasm (30). Many NF-κB-activating agents enable I-kappa- B (IKBKA or IKBKB) kinases to phosphorylate serine residues on the NFKBIα protein, thereby causing degradation via ubiquitination (31). Transcriptional products of NF-κB include tumour necrosis factor alpha (TNF-α), which plays a central role in psoriasis or PsA pathophysiology (32). rs12883343 is located at the 3’ of the NFKBIA gene. Although we found no evidence that this SNP is a cis-eQTL for NFKBIA mRNA levels in skin tissue, it could be regarded as a regulatory element in many cell types relevant to PsC and PsA. These include keratinocytes, Th 17 cells, neutrophils, CD14+ monocytes, fibroblasts, and osteoblasts (33). This may provide us with a new method by which to predict the risk of developing PsA and to acquire evidence indicating that PsA and PsC are genetically different. This is different from the results of previous studies. Explanations for this difference could include the presence of heterogeneous associations within different populations, or a lack of power in moderately sized case-control cohorts.
We also found a PsA-specific variant (rs 4921485) in the IL12B region, which appeared to be borderline evidence for PsA association (p = 4.50 × 10–3, OR 0.767), with no detectable association with PsC (p = 0.728, OR 0.969). It is the first time that this variant and its associated findings relative to PsA and PsC have been reported. Our results replicate the previously described association (Table V) of 5 alleles, which showed borderline evidence at a nominal p-value threshold of 0.05, with the risk of developing PsA. Four variants in IL23R and IL12B were included. This reveals the important role of the IL-12/IL-23 pathway in the pathogenesis of PsA and provides an explanation for the effective treatment of this condition using ustekinumab; a monoclonal antibody against IL-12 and IL-23 (34).
Table V. Single-nucleotide polymorphisms (SNPs) in previous studies
This study supports the hypothesis that there are differences in the underlying genetic architecture between PsA and PsC. The study was based on a limited number of PsA, PsC and control subjects from the Chinese population. Although we gave a more stringent definition of PsC to decreasing the patients who will go on to develop PsA, the PsC subset may still include patients who will ultimately develop PsA. Therefore, more studies that include a large number of affected case subjects are needed to detect or confirm the differential association between PsA and PsC. In parallel, studies investigating a large number of SNPs, GWAS in large sample sets, meta-analyses of genome-wide scan results, and large-scale analyses of rarer variants are needed to identify additional susceptibility loci in the Chinese population.
The current study found a previously unreported variant in NFKBIA, showing that a significant difference exists between PsA and PsC. The results contribute towards understanding the genetic aetiology of PsC in the Chinese population, as well as the differences in the genetic architecture of PsA and PsC. Further research is needed to identify the biological mechanisms that are most relevant to each clinical subtype, and to provide new targets for treatment.
The authors would like to thank the patients and doctors who supported and participated in this study.
Funding: This work was supported by the Innovation Project of Shandong Academy of Medical Sciences; and the Natural Science Foundation of Shandong Province (JQ201616, BS2013YY010).
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize Click to show fullsize
Click to show fullsize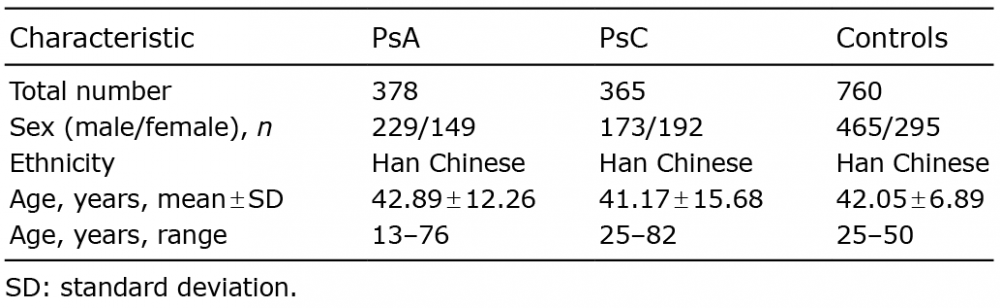 Click to show fullsize
Click to show fullsize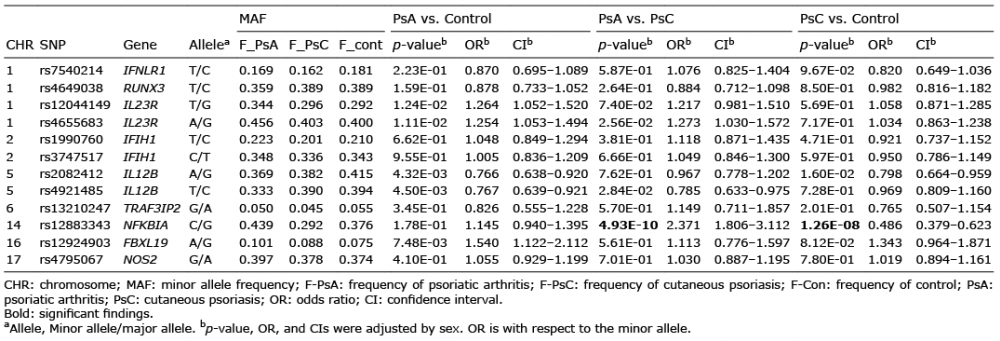 Click to show fullsize
Click to show fullsize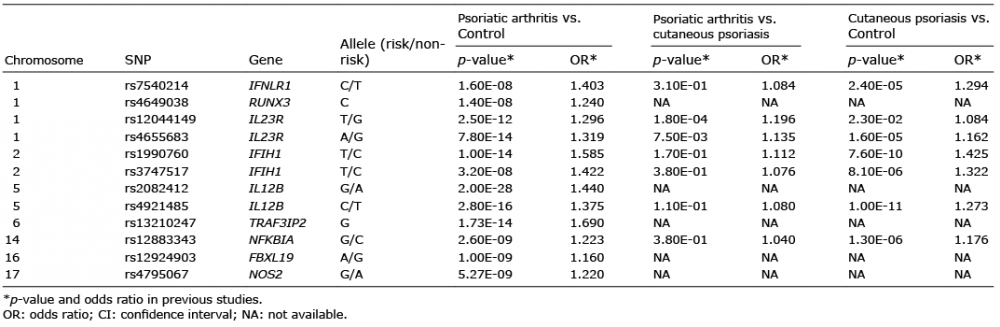 Click to show fullsize
Click to show fullsize