


Departments of 1Dermatology and Venereology, 3Physiology and 4Pathophysiology, University of Tartu, Raja 31, EE-50417 Tartu, 2Clinic of Dermatology, Tartu University Hospital, and 5Department of Reproductive Biology, Estonian University of Life Sciences, Tartu, Estonia. E-mail: tanel.traks@ut.ee
Accepted Feb 5, 2019; E-published Feb 6, 2019
Psoriasis is a non-contageous chronic inflammatory disease that predominantly affects the skin and joints. The most common clinical type is plaque psoriasis, characterized by well-demarcated and erythematous plaques with silvery scales (1). In addition to the effects on the skin, psoriasis is also associated with systemic inflammation and has frequent comorbidities, such as cardiovascular disease, diabetes and various cancers (1, 2). As a multifactorial disease, it has many potential environmental triggers, while the important contribution of genetic factors has been firmly established and extensively studied (3).
The corticotrophin-releasing hormone-proopiomelanocortin (CRH-POMC) system is organized as a cutaneous equivalent to the hypothalamic-pituitary-adrenal (HPA) axis and regulates local stress responses and melanogenesis in the skin (4). CRH and its receptor CRH-R1 that function as principal components of the HPA axis are also expressed in the skin (5). The binding of CRH to this receptor causes the synthesis of POMC in melanocytes and fibroblasts and the subsequent production of POMC-derived peptides (6). Overall, the POMC system includes its cleavage products α-, β-, and γ-melanocyte stimulating hormone (α-, β-, γ-MSH) and adrenocorticotropic hormone (ACTH); 5 melanocortin receptors (MC1R-5R); 2 endogenous melanocortin receptor antagonists: agouti signaling protein (ASIP) and agouti related neuropeptide (AgRP). The immunomodulatory effects of CRH-POMC can be both pro- and anti-inflammatory and therefore imbalances in its regulation would have implications for the inflammatory skin disorders (7).
In previous reports, concerning CRH-POMC expression in psoriasis, CRH, CRH-R1, POMC, MC2R, MC3R, MC4R and melanin-concentrating hormone receptor 1 (MCH-R1) were elevated in psoriasis patients, whereas the levels of ASIP, tyrosinase (TYR) and tyrosinase related protein (TYRP1) were reduced (8, 9). Since these expressional patterns, along with other functions, could be influenced by genetic factors, the current aim was to investigate the possible genetic associations between CRH-POMC and related genes and plaque psoriasis.
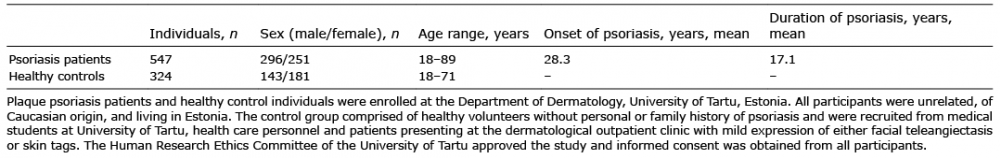
In total, 38 single nucleotide polymorphisms (SNPs) were selected from CRH, POMC, MC1R, MC2R, MC3R, MC4R, MC5R, ASIP, AGRP, TYR and dopachrome dautomerase (DCT) genes to be genotyped in an Estonian case-control sample (Table I). They intended to evenly cover each locus with the non-synonymous SNPs preferred, if available. SNPlex™ platform was used for genoytyping (Applied Biosystems, Foster City, California, USA) and Haploview v4.2 program was used for; 1) Hardy–Weinberg equilibrium evaluation, 2) allelic and haplotype association calculations and 3) permutation testing (10). The Solid Spine of LD algorithm integrated in Haploview v4.2 was applied in order to define the haplotype blocks. Differences in allele or haplotype frequencies between cases and controls were assessed by the chi-square test. The statistical significance threshold (p-value) was set to 0.05 for all tests. Ten thousand permutations were performed to correct p-values for errors of multiple testing.
Table I. Characteristics of study participants
All 38 SNPs were successfully genotyped and allele distributions in controls met the inclusion criteria for the Hardy-Weinberg equilibrium.
The most significant association involved the SNP rs6567166 located near the 3’ end of MC4R gene and the result withstood the correction for multiple testing (permutation adjusted p-value (padj) = 0.0236, risk allele odds ratio 5.45, 95% confidence interval 1.91−15.58; Table II). There are no previous reports discussing this specific polymorphism, but several other SNPs of this gene have been repeatedly associated with obesity and shown to affect different aspects of the MC4R, e.g. its transport, binding and signaling properties (11). It has been observed that obesity is twice as prevalent among psoriasis patients as in the general population (2) and although it is currently purely hypothetical, one could propose that mechanisms leading to both are intertwined and involve the MC4R as an important contributor. A recent study also examined SNP rs17782313 in the area of the MC4R in relation to psoriasis and while there was no association with disease severity, it was found to influence the incidence of arthritis, diabetes mellitus and obesity among patients (12). Further indication of the role of MC4R came from our previous study in which MC4R mRNA was elevated in lesional and non-lesional skin of psoriasis patients (9). Moreover, percutaneous treatment with α-MSH containing gel has been shown to decrease psoriasis symptoms (13) and despite MC4R not being its strongest affinity receptor, it could have a certain role in this process. We recently also published the results of our vitiligo genetic association study concerning CRH-POMC system genes (14). The control cohort largely overlapped with the present report and the same allele C of rs6567166 was significanly elevated among the patients. Summing up, rs6567166 may have an effect on MC4R functioning, or be in linkage disequillibrium with polymorphisms that do affect its function, and thereby influence psoriasis pathology.
Table II. Results of allelic association analysis
The rest of the obtained genetic associations did not remain significant after correction for multiple testing was applied. Still, the strongest among them was rs594647 from TYR gene. This SNP formed a haplotype block with 5 others (rs1042602, rs12791412, rs2000553, rs7358418 and rs10830250) and its haplotype AGATTC produced an association (p = 0.0024). It is interesting to note that another SNP, rs17793678, located just 33 bp from rs594647 was previously associated with malignant melanoma (15). Additionally, two intronic SNPs from DCT, one from MC2R and one from ASIP genes were nominally associated. Considering our mRNA expression results mentioned above, these findings could potentially reflect the impact of genetic factors on altered levels of TYR, MC2R and ASIP in psoriasis (9).
In conclusion, the results presented here may indicate true causal mechanisms underlying the pathogensis of plaque psoriasis, but should be considered preliminary due to the moderate sample size and small number of SNPs tested and would thus have to be confirmed through more extensive research. Until then, they are supported by the clear psoriasis phenotype of tested individuals, theoretical and empirical foundation of CRH-POMC role in this disease and statistical significance of the findings.
This work was supported by personal research grant [PUT1465] from the Estonian Research Council and institutional research grant [IUT20-46] from the Estonian Ministry of Education and Research.
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize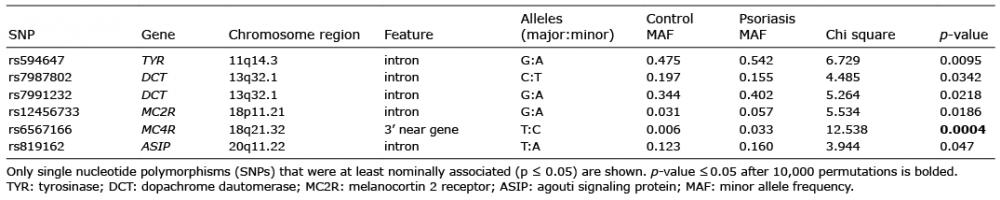 Click to show fullsize
Click to show fullsize