


Departments of 1Biochemistry, 2Microbiology and 4Pediatrics, College of Medicine, Ewha Womans University, 3Department of Health Sciences and Technology, Samsung Advanced Institute for Health Sciences and Technology, Graduate School, Sungkyunkwan University, Seoul, and 5Department of Biochemistry, College of Medicine, Gachon University, Incheon, South Korea
#These authors contributed equally to this manuscript.
Sphingosine-1-phosphate (S1P) is a signalling sphingo-lipid metabolite that regulates important cell processes, including cell proliferation and apoptosis. Circulating S1P levels have been reported to be increased in patients with psoriasis relative to healthy patients. The aim of this study was to examine the potency of S1P inhibition using an imiquimod-induced psoriasis mouse model. Both topical ceramidase and sphingosine kinase 1/2 inhibition, which blocks S1P generation, alleviated imiquimod-induced skin lesions and reduced the serum interleukin 17-A levels induced by application of imiquimod. These treatments also normalized skin mRNA levels of genes associated with inflammation and keratinocyte differentiation. Inhibition of sphingosine kinase 2, but not sphingosine kinase 1, diminished levels of suppressor of cytokine signalling 1 and blocked T helper type 17 differentiation of naïve CD4+ T cells; imiquimod-induced psoriasis-like skin symptoms were also ameliorated. These results indicate the distinct effects of sphingosine kinase 1 and sphingosine kinase 2 inhibition on T helper type 17 generation and suggest molecules that inhibit S1P formation, including ceramidase and sphingosine kinase 2 inhibitors, as novel therapeutic candidates for psoriasis.
Key words: psoriasis; CD4+ T lymphocyte; sphingosine kinase; sphingosine-1-phosphate; Th17 differentiation.
Accepted Mar 4, 2019; E-published Mar 5, 2019
Acta Derm Venereol 2019; 99: XX–XX.
Corr: Joo-Won Park, Department of Biochemistry, College of Medicine, Ewha Womans University, Seoul 07985, South Korea. E-mail: joowon.park@ewha.ac.kr; and Woo-Jae Park, Department of Biochemistry, College of Medicine, Gachon University, Incheon 21999, South Korea. E-mail: ooze@gachon.ac.kr
Sphingosine-1-phosphate is a bioactive lipid molecule involved in various cellular processes, such as cell proliferation and death. It can be formed by enzymes called sphingosine kinases. Elevation of sphingosine-1-phosphate in patients with psoriasis has been reported recently. Modulation of sphingolipids, including sphingsosine-1-phosphate, has emerged as a potentially powerful new clinical approach, and considerable progress has been made recently. Therefore, the effects of sphingosine-1-phosphate and its modulation on psoriasis should be assessed to uncover new therapeutic candidates for psoriasis. This study suggests molecules that inhibit sphingosine-1-phosphate formation, including ceramidase and sphingosine kinase 2 inhibitors, as novel therapeutic agents for psoriasis.
Psoriasis is a common, chronic immune-mediated inflammatory skin disorder (1). The important role of T-helper 17 (Th17) cells in the development of psoriasis lesions has been examined in many studies (2, 3). In patients with psoriasis, this unique CD4+ T-cell subset, characterized by interleukin-17 (IL-17) production, is found in the dermis of psoriasis lesions; the levels of circulating Th17 cells are also elevated (3, 4). IL-17A and IL-22 participate in the in?ammatory process during psoriasis by promoting the accumulation of T cells and neutrophils in the psoriatic lesion (5). Therapeutic approaches that modulate Th17 cells or the effector cytokines they produce have shown some efficacy for the treatment of psoriasis (5).
Sphingolipids are a complex group of lipids that contain the long-chain bases sphinganine (dihydrosphingosine) and sphingosine. Attaching fatty acyl CoA to a long-chain base via an amide linkage generates ceramide (Fig. S1), which can be modified further to produce more complex sphingolipids (e.g. sphingomyelins, cerebrosides, and gangliosides) (6). Sphingolipids have structural roles in lipid membranes and can also serve as sources of signalling molecules (7, 8). Signalling pathways affected by sphingolipids and their derivatives include those associated with apoptosis, proliferation, inflammation, and mitochondrial function (7, 8). Sphingosine-1-phosphate (S1P) is a bioactive sphingolipid metabolite involved in many critical cell processes (8). It can be formed by sphingosine kinases (SKs), which catalyse the phosphorylation of sphingosine. There are 2 types of SKs; SK1 is in the cytosol and plasma membrane and SK2 is in the nucleus (9). Although SK1 and SK2 share 80% similarity and 45% overall sequence identity, each has a distinct distribution, regulation, and pathophysiological function in cells (10). S1P generated by SK1 can be exported out of cells via transporters, such as spinster homolog 2. S1P regulates various processes by binding to specific G protein-binding receptors (i.e. S1P receptors (S1PRs) 1–5) and mainly acts as a pro-survival and pro-proliferative molecule (11). In contrast, studies of SK2 found that its overexpression induces cell cycle arrest and apoptosis (10). Nuclear S1P produced by SK2 inhibits histone deacetylase 1/2 activity and acts as an epigenetic and transcriptional regulator (10, 12). S1P also has an important role in immunological function because the S1P concentration gradient between various tissues promotes S1PR1-dependent migration of T cells from secondary lymphoid organs into the lymphatic and blood circulation (11).
Results of human studies indicate that there is an association between S1P and psoriasis. Plasma levels of sphingoid bases (e.g. S1P) are elevated in severely, but not in mildly, affected patients with psoriasis, relative to healthy controls (13, 14). Effects of modulating S1PR1 in Th17 differentiation have been examined in experimental autoimmune encephalomyelitis animal model studies. Deletion of S1PR1 in Th17 cells confers resistance to experimental autoimmune encephalomyelitis (15). FTY720, a functional antagonist for S1PR1, attenuates Th17 differentiation by downregulating STAT3 phosphorylation when FTY720 is exposed before establishment of the Th17 phenotype (16). The numbers and activities of Th17 cells are augmented in double transgenic mice expressing OTII TCRs specific for ovalbumin peptide 323–339 and a high level of transgenic S1PR1 upon OVA stimulation (17). However, the role of S1P and the associated mechanisms in patients with psoriasis remain to be fully described. The objective of this study was to investigate the effects of topical S1P modulation in psoriasis-like skin disease. It was found that S1P generated by SK2 has a critical role in Th17 differentiation via regulation of suppressor of cytokine signalling 1 (SOCS1).
All materials were purchased from Sigma Aldrich (St Louis, MO, USA) except: (i) imiquimod (IMQ) cream (5%, Aldara; Dong-Ah Pharmaceutical, Seoul, South Korea); (ii) Ceranib-1, MP-A08, and CYM50358 (Tocris Bioscience, Bristol, UK); (iii) Ceranib-2, PF-543, ABC294640, JTE013, and CAY10444 (Cayman Chemical, Ann Arbor, Michigan, USA), 4) anti-Ki67, anti-SK1, and anti-SK2 antibodies (Abcam, Cambridge, UK).
All mouse experimental procedures were approved by the Animal Care and Use Committee of the Ewha Womans University School of Medicine (ESM-17-0388). C57BL/6 mice (male, 18–20 g, 8 weeks of age) were purchased from OrientBio (Seongnam, South Korea) and housed under specific-pathogen-free conditions on a 12 h light:12 h dark cycle with free access to food and water. To induce psoriasis-like skin inflammation, mice were treated once daily with a topical dose of 83 mg (18) of IMQ cream applied to the shaved back region for 6 consecutive days, as described previously (19). Normal control mice were treated similarly with a control vehicle cream (Vaseline Lanette cream, Fagron, Rotterdam, The Netherlands). For prophylactic chemical treatment, mice were exposed once daily to topically administered MP-A08, Ceranib-2, PF-543, or ABC294640. MP-A08, Ceranib-2, and ABC294640 were used at 25 µg/40 µl surfactant (100% ethanol, propylene glycol, and H2O (EPH) at 2:1:1 (v/v/v)); PF-543 was used at 12.5 µg/40 µl surfactant applied to the shaved back skin. Separate control groups received the vehicle control (EPH) for 3 consecutive days before inducing skin inflammation.
The severity of inflammation of the back skin was monitored and graded using the clinical Psoriasis Area and Severity Index (PASI), as previously described (18). Erythema, scaling, and thickening were scored independently on a scale from 0 to 4 (0, none; 1, slight; 2, moderate; 3, marked; 4, very marked). The cumulative score represented the severity of skin inflammation.
The specimens from mouse back skins were fixed using 4% (w/v) paraformaldehyde, embedded in paraffin, and then sectioned at a thickness of 5 µm. Slides were stained using haematoxylin and eosin (H&E) according to standard methods for histological evaluation.
For the immunohistochemistry, the sections from paraffin block were deparaffinized using xylene and rehydrated with a graded series of alcohols. Endogenous peroxidase activity was quenched using 0.3% (v/v) hydrogen peroxide and 0.5% (v/v) HCl in methanol. Antigen retrieval was performed by heating the slides in 10 mM citrate buffer (pH 6.0). The slides were pre-incubated with normal goat serum (10%) for 1 h at room temperature to block non-specific reagent binding. Overnight incubation at 4°C with anti-Ki67 was followed by incubation with horseradish peroxidase (HRP)-conjugated secondary antibody. Staining was detected using the Vectastain Elite ABC Kit (Vector Laboratories, Burlingame, CA, USA), and 3,3?diaminobenzidine (DAB) was used as the chromogen. The slides were counterstained using haematoxylin.
Splenic naïve CD4+ T cells were cultured in Th17 polarizing conditions and then re-stimulated with coated anti-mouse CD3e mAb (1 µg/ml) (BD Biosciences PharMingen, San Diego, CA, USA) and soluble anti-mouse CD28 mAb (2 µg/ml) (BD Biosciences PharMingen) for 40 h. The mice were humanely euthanized and mouse serum was obtained at day 6 of IMQ treatment. Mouse serum and culture supernatant IL-17A levels were determined using the LEGEND MAX™ mouse IL-17A enzyme-linked immunosorbent assay (ELISA) kit with pre-coated plates (BioLegend, Inc, San Diego, CA, USA), according to the manufacturer’s instructions.
Total mRNA was isolated from the skin samples using RNeasy Mini Kit (Qiagen, Valencia, CA, USA). Complementary DNA was synthesized using ReverTraAce qPCR RT master mix with gDNA remover (Toyobo, Osaka, Japan), according to the manufacturer’s instructions. Real-time PCR was performed using the SYBR Green PCR Master Mix (Applied Biosystems, Warrington, UK) and an ABI PRISM 7500 sequence detection system (Applied Biosystems). All gene expressions were normalized to the GAPDH reference gene. Relative gene expression was determined as 2−ΔΔCt, as previously described (20). The primers used are shown in Table SI.
Naïve CD4+ T cells were purified from mouse spleen samples using magnetic beads (Naive CD4+ T Cell Isolation Kit, mouse; Miltenyi Biotec, Gladbach, Germany). Naïve CD4+ T cells were primed with plate-bound anti-CD3 (1 µg/ml) (BD Biosciences PharMingen, San Diego, CA, USA) and soluble anti-mouse CD28 (2 µg/ml) (BD Biosciences PharMingen) in round bottomed 96-well plates with RPMI1640 medium containing a 10% foetal bovine serum and 1% penicillin/streptomycin solution (Hyclone, Logan, UT, USA). For Th17 differentiation, the cells were cultured in the presence of soluble anti-CD28 (1 μg/ml), rmIL-2 (20 ng/ml; Peprotech, London, UK), rmTGF-β (5 ng/ml; R&D Systems, Minneapolis, MN, USA), rmIL-6 (25 ng/ml; Peprotech), and anti-IL-4 and anti-IFN-γ neutralizing antibodies (10 μg/ml, Biolegend) for 5 days. The cells were then washed with PBS, transferred to a new plate to incubate for 2 days, and were then re-stimulated for 40 h with Dynabeads Mouse T-Activator CD3/CD28 (Invitrogen, Carlsbad, CA, USA). For the chemical inhibition, the following chemicals were then added to the sample medium: Ceranib-1 (30 μM; (21)), Ceranib-2 (30 μM; (21)), MP-A08 (15 μM; (22)), PF-543 (30 nM; (23)), ABC294690 (26 μM; (24)), JTE013 (5 μM; (25)), CAY10444 (5 μM; (25)), and CYM50358 (5 μM; (25)).
Skin protein samples (50 μg) were run on sodium dodecyl sulphate (SDS)-polyacrylamide gel (10% (w/v)) electrophoresis and then transferred onto nitrocellulose membranes. The membranes were incubated with primary antibodies overnight at 4°C, followed by peroxidase-conjugated secondary antibodies (Jackson ImmunoResearch, West Grove, PA, USA) for 1 h at room temperature after blocking with 5% (w/v) bovine serum albumin (BSA) for 1 h. The bands were detected by Bio-Imaging Analyzer LAS-4000 (Fuji, Tokyo, Japan) using SuperSignal West Pico chemiluminescent substrate (Thermo Fisher Scientific).
SK1 and SK2 activities were measured as previously described (26), but modified. Skin tissues were homogenized and sonicated in lysis buffer (50 mM Tris/HCl buffer (pH 7.4), 150 mM NaCl, 10% glycerol, 0.05% Triton X-100, 1 mM dithiothreitol, 2 mM Na3VO4, 10 mM NaF, 10 mM β-glycerophosphate, 1 mM EDTA) and 50 µg of protein was used for each activity assay. Different incubation buffers were used for SK isoform as described previously (26). 100 µmol/l NBD-sphingosine with serum-free BSA (4 mg/ml) was added and incubated at 37°C in 30 min. Synthesized S1P was extracted and separated by thin-layer chromatography (TLC) using a solvent system containing 1-butanol:ethanol:acetic acid:water (80:20:10:20 [v/v/v/v]). S1P on TLC were identified via Typhoon 9410 scanner (GE Amersham Biosciences, Pittsburgh, PA, USA) and bands were quantified using ImageQuant software (GE Amersham Biosciences).
The results were presented as means ± standard error of the mean (SEM) values. Statistical significance was analysed using the Student’s t-test, 1-way analysis of variance (ANOVA), or 2-way ANOVA. GraphPad PRISM 6 statistical software (GraphPad Software) was used for the analysis. A p-value < 0.05 was considered to indicate a statistically significant result.
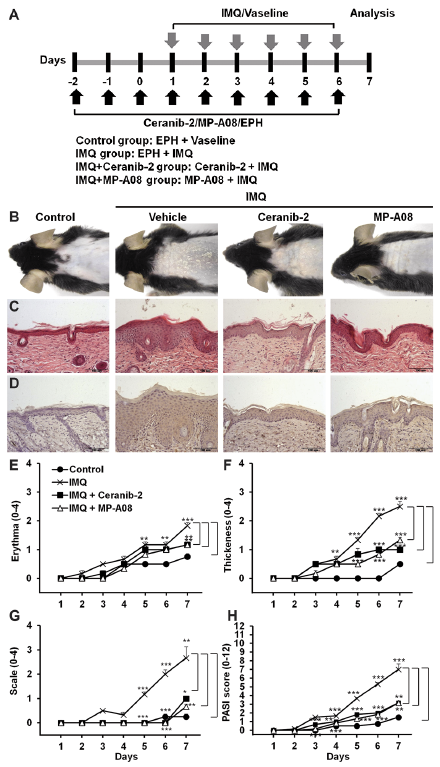
Based on the reports that circulating S1P levels are increased in individuals with psoriasis (13, 14), the current study investigated the effects of topical S1P modification in psoriasis-like skin disease. Psoriasis-like skin inflammation was induced by applying IMQ (a toll-like receptor 7/8 agonist) to the back skin of the mice (19). To investigate whether blocking S1P production can affect the severity of psoriasis-like skin inflammation, Ceranib-2, a ceramidase inhibitor, or MP-A08, a SK1/2 inhibitor, were applied topically as a pre-treatment for 3 days to reduce S1P levels. IMQ was then applied with the inhibitors to the same area, for 6 consecutive days (Fig. 1A). Consistent with the result of a previous study (19), IMQ-treated skin lesions had psoriasis-like changes in the epidermis (e.g. hyperplasia and erythema) (Fig. 1B). The H&E-stained skin sections revealed that pre-treatment with Ceranib-2 or MP-A08 reduced the severity of IMQ-induced erythema and scaling (Fig. 1B), and reduced epidermal thickness (Fig. 1C). Ki-67 is a nuclear protein associated with cellular proliferation. To confirm the proliferative response in the epidermis, Ki-67 was examined using an immunohistochemical method. Epidermal Ki-67 immunoreactivity was significantly higher in the IMQ-treated groups compared with normal epidermis. However, treatment with Ceranib-2 or MP-A08 reduced Ki-67 immunohistochemical staining in the IMQ-treated groups (Fig. 1D).
To evaluate daily skin changes after IMQ application, grades of erythema, scaling, and thickening was evaluated using the PASI score (Fig. 1E–H). On the third day of IMQ application, the mean PASI score for the IMQ-treated mice started to increase; on the seventh day of IMQ application, the mean ± SEM PASI score for this group was 7 ± 0.63. In contrast, on the seventh day of IMQ application, the PASI scores for the Ceranib-2- and MP-A08-treated groups were 3.17 ± 0.26 and 3.17 ± 0.26, respectively. The differences between the Ceranib-2- or the MP-A08-treated IMQ group and the Vaseline®-treated IMQ group were statistically significant. These results revealed the protective effect of S1P reduction in psoriasis-like skin disease.
Fig. 1. Imiquimod (IMQ)-induced skin lesions are alleviated by topical application of Ceranib-2 or MP-A08. (A) Study plan: C57BL/6 mice were treated daily with IMQ cream or the control vehicle cream on the shaved back for 6 consecutive days. To inhibit sphingosine-1-phosphate (S1P) generation, Ceranib-2 (a ceramidase inhibitor) or MP-A08 (a sphingosine kinase 1/2 inhibitor) were applied at 3 days before application of the IMQ cream and were maintained during treatment with IMQ. (B) Phenotypical presentation of mouse back skin after 6 days of treatment. (C) Haematoxylin and eosin-stained skin tissue sections. (D) Analysis of Ki-67 expression in paraffin-embedded skin tissue sections. The severity of (E) erythema, (F) thickening and (G) scaling were scored using the clinical Psoriasis Area and Severity Index (PASI), as previously described (20). (H) The cumulative PASI score represents the severity of skin inflammation. Results are presented as mean ± SEM values (n = 5). **p < 0.01, ***p < 0.001. The images are representative of 3 independent experiments (magnification ×20).
The immune-mediated response, including Th17 induction, is one of the important mechanisms that contribute to psoriasis severity. To determine whether topical use of Ceranib-2- or MP-A08 decreased the systemic immune response induced by IMQ, we compared the size and cellularity of lymphoid organs. Following 7 days of IMQ application, the results indicated that there was a consistent statistically significant spleen and lymph node enlargement in Vaseline®-treated group (Fig. 2A, B). Consistent with the improvement in skin lesions upon topical use of Ceranib-2- or MP-A08 (Fig. 1), the inguinal lymph node and spleen enlargement in the IMQ-treated mice was smaller in the Ceranib-2- and MP-A08-treated groups compared with the EPH-treated group (Fig. 2A, B). Both the mean cell numbers in the inguinal lymph nodes and the mean spleen weights were significantly reduced in the Ceranib-2- and MP-A08-treated groups, compared with the EPH-treated group (Fig. 2C, D). The mean numbers of splenocytes were reduced in the Ceranib-2-treated group, compared with the EPH-treated group (Fig. 2E). Because IL-17A and the Th17 pathways have critical roles in the pathology of psoriasis (5), we examined IL-17A levels in blood. IMQ application markedly increased serum IL-17A levels; Ceranib-2 or MP-A08 treatment significantly reduced serum IL-17A levels (Fig. S2A). These results indicated that topical inhibition of S1P generation hampers IMQ-induced systemic immune responses, including Th17 induction.
Fig. 2. Inhibition of sphingosine-1-phosphate (S1P) generation reduces the enlargement of lymphoid organs upon treatment with imiquimod (IMQ). Gross morphological examination of (A) inguinal lymph node and (B) spleen. (C) Total cell numbers in inguinal lymph nodes were counted in each group. (D) Spleen weights and (E) total cell numbers were investigated. Results are mean ± SEM values (n=5). *p < 0.05, ***p < 0.001.
Next it was examined whether Ceranib-2 or MP-A08 treatment can directly alter pro-inflammatory cytokine gene expression in the skin lesion. Similar to the serum results (Fig. S2A), mRNA levels of pro-inflammatory cytokines, including IL-17A, IL-17F, and TNF-α, were elevated in IMQ-applied skin lesions. Topical Ceranib-2 or MP-A08 treatment effectively normalized these alterations in gene expression (Fig. S2B–D). Keratins K6 and K16 are normally absent from the interfollicular epidermis. However, they are induced when a hyperproliferative stimulus, such as IMQ application, is present. Consequently, they are used as markers of keratinocyte hyperproliferation (27). IMQ application to the skin resulted in increased expression of K6 and K16 keratin genes; Ceranib-2 or MP-A08 treatment significantly suppressed expression of these genes (Fig. S2E, F). These results indicated that inhibiting S1P formation directly mitigated the regional pro-inflammatory immune response and reduced the keratinocyte hyperproliferation caused by application of IMQ. Ceramidase converts ceramide into sphingosine. Therefore, ceramidase inhibition elevates ceramide and diminishes sphingosine and its derivative S1P (21). Because SK inhibition also reduces S1P levels, the current study examined how S1P reduction alleviates the psoriasis-like skin phenotype.
Systemic and regional IL-17A levels were lower as a consequence of Ceranib-2 or MP-A08 treatment (Fig. S2A, B). Therefore, the current study investigated whether inhibiting S1P generation would directly affect Th17 differentiation of naïve CD4+ T cells. Both SK1/2 inhibition with MP-A08 and ceramidase inhibition using Ceranib-1 or Ceranib-2 significantly reduced IL-17A levels secreted from differentiated CD4+ T cells (Fig. S3A). These results indicated that these treatments inhibited Th17 differentiation of naïve CD4+ T cells. A specific SK1 inhibitor (PF-543 (23)) and a specific SK2 inhibitor (ABC294640 (21, 24)) were then used to determine which SK isotype was important for S1P generation involved in Th17 differentiation of naïve CD4+ T cells. Kinase selectivity of PF-543 and ABC294640 has been tested in the previous studies (23, 24). Only SK2 inhibition negatively affected Th17 differentiation of naïve CD4+ T cells (Fig. S3B). S1P generated via SK1 can be exported out of the cells and exert its action through the S1PRs, S1PR1–5 (11). Because S1PR2–4 were the S1PRs mainly expressed in murine CD4+ T cells (Fig. S3C), we also investigated whether inhibition of S1PR2–4 reduced Th17 differentiation of naïve CD4+ T cells. JTE013, CAY10444 and CYM50358 are specific antagonists of S1PR2, 3 and 4, respectively (25). In accordance with SK1 inhibition, treatment with JTE013, CAY10444 and CYM50358 did not affect Th17 differentiation of naïve CD4+ T cells (Fig. S3D–F). This study also examined SOCS1 and 3 levels, because SOCS1 and 3 are essential regulators for helper T-cell differentiation (28, 29). Th17 differentiation is strongly suppressed in SOCS1 deficient naïve CD4+ T cells or in SOCS3 overexpressed T cells (28). In accordance with suppression of Th17 differentiation upon SK2 inhibition, SK2 inhibition (but not SK1 inhibition) resulted in a statistically significant reduction in SOCS1 mRNA levels (Fig. S3G). However, SOCS3 transcription was not altered after SK1 or SK2 inhibition (Fig. S3H). These results indicated that S1P generation produced by SK2, but not SK1, was necessary for Th17 differentiation of naïve CD4+ T cells. Blocking the S1P generation produced by SK2-attenuated generation of Th17 cells from naïve CD4+ T cells.
This study also examined whether SK2 inhibition alleviated psoriasis-like skin disease in vivo (Fig. 3A). IMQ treatment increased not only protein expressions of SK1 and SK2, but also kinase activities of SK1 and SK2 (Fig. S4A, B). Topical application of PF543 and ABC294640 specifically reduced SK1 and SK2 activity, respectively (Fig. S4B), with no significant alteration of protein expression (Fig. S4A). Examination of the H&E-stained skin sections revealed that SK2 inhibition using ABC294640, but not SK1 inhibition using PF-543, reduced IMQ-induced erythema and scaling (Fig. 3B), and epidermal thickness (Fig. 3C). The PASI score of the SK2-inhibited group was also markedly reduced compared with that of the EPH-treated IMQ group (Fig. 3D–G). To determine whether SK2 inhibition decreased the systemic immune response induced by IMQ, the current study compared lymphoid organ size and cellularity. Consistent with the improvement in skin changes after topical SK2 inhibition (Fig. 3), the sizes of the inguinal lymph nodes and spleen in IMQ-treated mice were smaller in the ABC294640-treated group compared with the EPH-treated IMQ group (Fig. S5A, B). Both the mean cell numbers in the inguinal lymph nodes and the mean spleen weights were significantly diminished in the ABC294640-treated group compared with the EPH-treated IMQ group (Fig. S5C, D). Splenocyte cell numbers were also decreased in the ABC294640-treated group compared with the EPH-treated IMQ group (Fig. S5E).
Fig. 3. Sphingosine kinase (SK)2 inhibition results in protective effects in psoriasis-like skin disease. (A) Study plan: C57BL/6 mice were treated once daily with imiquimod (IMQ) cream or control vehicle cream applied to the shaved back for 6 consecutive days. Once daily PF-543 (a SK1 inhibitor) and ABC294640 (a SK2 inhibitor) application began at 3 days before application of IMQ cream and maintained during IMQ treatment. (B) Phenotypical presentation of mouse back skin after 6 days of treatment. (C) Haematoxylin and eosin-stained skin tissue sections. (D) Erythema, (E) thickening and (F) scaling were scored independently on a scale from 0 to 4 using the Psoriasis Area and Severity Index (PASI). (G) The cumulative PASI score represented the severity of skin inflammation. Results are mean ± SEM (n=5). *p < 0.05, **p < 0.01, ***p < 0.001. The images are representative of 3 independent experiments (magnification ×20).
Sphingolipids are a class of lipids characterized by 18-carbon amino-alcohol backbones that are synthesized in the endoplasmic reticulum from non-sphingolipid precursors (i.e. amino acids, serine, and palmitoyl CoA (30). Modification of this basic structure results in the vast family of sphingolipids that have critical roles in membrane biology and provide bioactive molecules that determine the fates of cells (30). Alterations in sphingolipid homeostasis are associated with the development of pathological disorders. Therefore, therapeutic strategies to modulate sphingolipid metabolism and their related signalling pathways have emerged rapidly (31). High levels of S1P, a bioactive signalling molecule formed by SK1 or SK2, have been found in patients with psoriasis (13, 14). Therefore, the current study investigated the effects of S1P modulation using an IMQ-induced mouse model of psoriasis. This study revealed an important role of S1P generated by SK2 in Th17 differentiation of naïve CD4+ T cells and in the pathogenesis of psoriasis.
Using an IMQ-induced psoriasis mouse model, it was found that topical inhibition of S1P formation alleviated psoriasis-like skin symptoms. However, in contrast to the results of this study, anti-proliferative and anti-inflammatory effects of S1P in mouse models of psoriasis have been reported previously (32). Schaper et al. found that topical administration of S1P diminishes IMQ-induced ear swelling and epidermal thickness in the ear; they suggested S1P as a new option for the treatment of mild to moderate psoriasis lesions (32). However, they found that topical application of S1P had no effect on the results of a histological evaluation of the back skin in the IMQ-induced psoriasis mouse model; the psoriasis severity score was also increased up to the third day of IMQ treatment (32). This result is consistent with the results of our study, which revealed the protective effect of S1P inhibition on psoriasis-like skin lesions. Schaper et al. found that IL-17 cytokine levels in ear homogenates increased after S1P application, but this result was not statistically significant (32). Liao et al.’s study also revealed the effectiveness of S1P to enhance the development of Th17 cells (33). These results are also consistent with the results of our study, which found that inhibition of S1P generation could hamper Th17 differentiation of naïve CD4+ T cells. Taken together, these results suggest that normalizing the elevated S1P levels that occur in patients with psoriasis represents a novel and effective therapeutic approach.
Psoriasis is among the most prevalent T-cell-mediated disorders. It was originally regarded as a Th1-mediated skin inflammation, but the pathogenesis of psoriasis is mainly due to the activity of Th17 cells (34). IL-17A has a pivotal role in maintaining inflammation in psoriatic plaques, and Th17 cells occur in psoriatic skin lesions at much higher numbers compared with healthy skin (34–36). Activated Th17 cells secrete IL-17A, IL-17F, IL-22 and IL-9, which enhance the inflammatory response of keratinocytes, and clinical trials with anti-IL-17 antibody find very rapid and highly effective outcomes in patients with cutaneous psoriasis (34). Therefore, regulating Th17 cell generation and activation can effectively modulate the pathogenesis of psoriasis. The current study found that inhibiting generation of S1P via SK2 efficiently blocked Th17 differentiation from naïve CD4+ T cells by decreasing SOCS1 levels. However, SK1 inhibition did not affect Th17 differentiation from naïve CD4+ T lymphocytes, or SOCS1 levels. SOCS1 is necessary for Th17 differentiation; it suppresses the antagonistic effect of IFN-γ on STAT3 and the SMADs pathway (29). The different role of SK1 and SK2 has been revealed using disease models (e.g. experimental colitis and cancer) (10). Compared with wildtype littermates, SK1 knockout mice treated with dextran sulphate sodium have a significantly alleviated colitis phenotype that includes less colon shortening and histologically apparent colon damage, and a less severe systemic inflammatory response (37). However, SK2 deficiency exacerbates colitis-associated tumorigenesis when SK2 knockout mice are injected with the colonotropic mutagen azoxymethane, followed by 3 rounds of oral administration of dextran sulphate sodium to trigger chronic inflammation (38). The tissue distributions and intracellular locations of SK1 and SK2 are also different (10). SK1 is localized in cytoplasm and can be re-localized to the plasma membrane to mediate pro-survival and pro-proliferative signalling. SK2 possesses nuclear localization and export signals; it inhibits DNA synthesis and regulates epigenetic modifications via interaction with and modulation of HDAC1/2 in the nucleus (10, 12). S1P generated by SK2 directly affects HDAC1/2 activity (12). The current study found that SK2 was the main SK isoform expressed in naïve CD4+ T cells (data not shown), and SK2 inhibition of naïve CD4+ T cells incubated in Th17 differentiating medium suppressed SOCS1 expression. The mechanism via which blocking S1P generation by SK2 suppressed SOCS1 transcription in CD4+ T cells remains to be determined, but modulation of HDAC1/2 by S1P is 1 possibility. Similar to S1P, Trichostatin A acts as a histone deacetylase inhibitor; it also leads to the upregulation of SOCS1 expression in colorectal cancer cells via hyperacetylation of histones associated with the SOCS1 promoter. Thus, the S1P increase in patients with psoriasis (13, 14) might induce the Th17 response by enhancing expression of SOCS1. Blocking S1P may become a novel therapeutic strategy for psoriasis.
FTY720, a functional antagonist for S1P receptor 1, was approved by the US Food and Drug Administration (FDA) as an oral medication to treat the relapsing form of multiple sclerosis. Since that approval, chemicals that modulate sphingolipids have been developed and are being evaluated in clinical trials. In summary, the results of this study suggested molecules that inhibit S1P formation, including ceramidase and SK2 inhibitors, as novel therapeutic candidates for psoriasis. S1P generation by SK2, but not by SK1, may have an important role in Th17 pathway-mediated immune diseases.
This work was supported by grants from the Health Technology R&D Project (HI14C2445) of the Ministry of Health and Welfare, Republic of Korea, from the National Research Foundation of Korea (NRF) grant funded by the Korea government (Ministry of Education) (NRF-2016R1D1A1B03935706), and by intramural research promotion grants from Ewha Womans University School of Medicine.


 Click to show fullsize
Click to show fullsize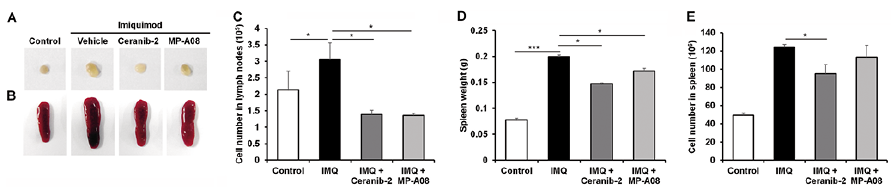 Click to show fullsize
Click to show fullsize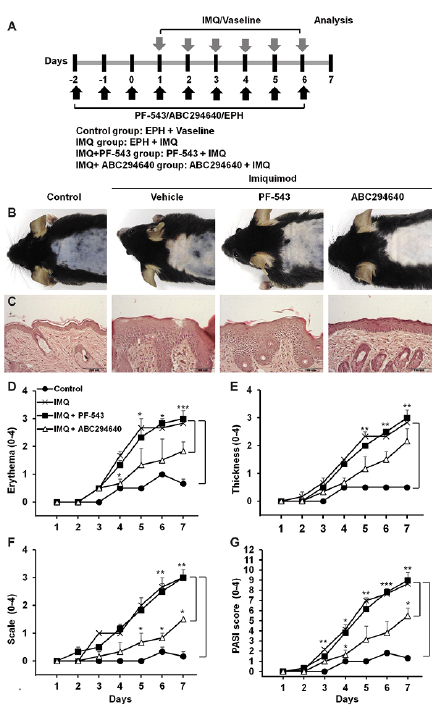 Click to show fullsize
Click to show fullsize